
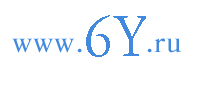

СТЕЛЯРНАЯ ТЕОРИЯ
При рассмотрении строения стебля и корня упоминалась стель, или осевой цилиндр, как область органа, включающая систему проводящих тканей и отграниченная от первичной коры специализированным, внутренним слоем последней - эндодермой, или крахмалоносным влагалищем. Корень в его строении консервативен, и конструкция его стели в основных чертах сходна у высших растений всех систематических групп различных геологических эпох. Строение же стели в стебле проходило в процессе эволюции (в филоморфогенезе) ряд этапов. Строение стели взрослых особей современных голосеменных и покрытосеменных представляет конечные ступени филоморфогенеза стели.
Рассмотрим в основных чертах последовательные этапы эволюции стели. Изложение поясним примерами строения ее у растений прошлых геологических эпох или у современных консервативных папоротникообразных, сохранивших стель более или менее примитивной конструкции. Предварительно напомним, что стель может включать, кроме проводящих пучков и арматурных тяжей или пластинок, еще и паренхиму. Паренхима, находящаяся в центре стели, представляет сердцевину (medula), паренхима между проводящими пучками образует первичные сердцевинные лучи, паренхима кнаружи от проводящих пучков - перицикл. Родоначальный, примитивный тип стели - протостель (рис. 181, А). Эта стель без сердцевинных лучей, состоящая из тяжа ксилемы и из окружающей его флоэмы.
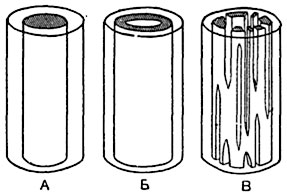
Рис. 181. Модели:
А - протостели; Б - сифоностели; В - диктиостели.
Протостелью обладали осевые органы, еще не расчлененные на стебли и корни, девонских псилофитов (Psilophyta). У наиболее примитивных из них (Rhynia, Hornea) вдоль органа проходил в центре тяж ксилемы, состоявший из кольчатых трахеид, более узких в центре. Флоэма, окружавшая ксилемный тяж, была построена из удлиненных тонкостенных клеток с наклонными поперечными перегородками без ситечек. Перицикл и эндодерма еще не дифференцировались. Стель резко выделялась на поперечных срезах от окружающей коровой паренхимы в силу малых размеров и тонкостенности клеток флоэмной части стели. Многие ископаемые папоротники и некоторые из современных (как Gleichenia) обладают протостелью. Размеры растений, обладающих протостелью, малы: диаметр протостели не превышает 0,3-3 мм.
209
У псилофитов, получивших в процессе эволюции более крупные размеры и дифференцировку надземного побега на стебель и листья, и у других папоротникообразных происходило параллельно с увеличением размеров их органов усложнение формы и строения стели. Морфолог Боуэр выдвигает, опираясь на основные положения учения Дарвина, физиологическое объяснение того явления, что в процессе эволюции под действием естественного отбора закреплялись изменения стели, выражающиеся в определенных чертах ее усложнения. Передача веществ из проводящих тканей в окружающие их живые паренхимные клетки и передвижение веществ в обратном направлении тем лучше, чем больше поверхность соприкосновения между проводящими и живыми тканями. По математическим законам, увеличение тела с сохранением геометрического подобия влечет за собой увеличение поверхностей пропорционально квадрату любого линейного размера, а увеличение объемов пропорционально кубу линейного измерения. Из этого следует, что если орган увеличивается, сохраняя неизменными пропорции в общих размерах и в размерах стели, то поверхность соприкосновения между проводящими тканями и живыми клетками органа становится физиологически недостаточной. Выходом из этого положения являлись происшедшие на базе действия естественного отбора усложнения в форме стели. Уже у более крупных и более высокоорганизованных псилофитов (у Asteroxylon) стель приобрела резко выраженную в поперечных сечениях лопастно-звездчатую форму - стала актиностелью.
У Asteroxylon узкие трахеиды протоксилемы расположены по углам стели (на концах лопастей звезды в поперечных сечениях). В промежутках между лучами ксилемы находятся флоэмные группы. Имеются уже намеки на появление эндодермы.
Актиностель найдена и у некоторых ископаемых папоротников со стелью диаметром до 15 мм.
Другим средством увеличения поверхности соприкосновения проводящих тканей с живой паренхимой является "витализация" ("оживление") стели. В стебле древних папоротникообразных - не только псилофитов, но и большинства видов лепидодендрона - ксилема состояла сплошь из трахеид. Из растений крупных размеров выживали те, стель которых изменялась в направлении витализации; часть клеток в ксилеме оставались живыми, паренхимными. Некоторые клетки в стели предков, у которых утолщались стенки, появлялись окаймленные поры и исчезало живое содержимое, в организме потомков останавливались в своем онтоморфогенезе на стадии живых клеток с тонкими стенками и простыми порами.
У ряда вымерших крупных папоротников (например, Asterochlaena) обнаружена промежуточная стадия: наличие "витализованной", или "смешанной", сердцевины - в центральной части ксилемы среди трахеид находились живые паренхимные клетки. У некоторых из сохранившихся доныне во флоре земного шара примитивных папоротников (У Ophioglossum, Βοtrychium) в молодом стебле имеется протостель с ксилемным тяжом, состоящим сплошь из трахеид. По мере роста стебля в длину продолжение тяжа приобретает паренхимную сердцевину, содержащую кое-где трахеиды.
Стель, содержащая в центре паренхимную сердцевину (так называемая сифоностель, рис. 181, Б), может отграничиваться от сердцевины внутренней эндодермой, дифференцирующейся в сердцевине (как папоротника Todea hymenophylla). У некоторых папоротников имеется и внутренняя флоэма - в виде неполного кольца (у папоротника Osmunda). У более высокостоящих папоротников выработалась сифоностель, имеющая сердцевину с внутренней эндодермой, кольцо внутренней флоэмы, кольцо ксилемы и наружное кольцо флоэмы, граничащее с наружной эндодермой. Стель подобного типа называется амфифлойной сифоностелью
210
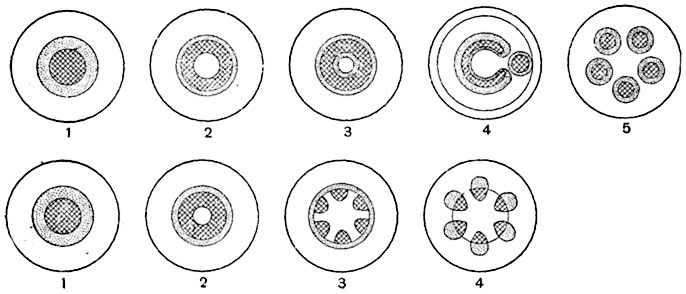
Рис. 182. Различные направления филоморфогенеза протостели.
Ксилема отмечена штриховкой, а флоэма - точками. Верхний ряд: 1 - протостель; 2 - эктофлойная сифоностель; 3 - амфифлойная снфоностель (соленостель); 4 - разрыв трубки соленостели, образование частной протостели; 5 - диктиостель. Нижний ряд: 1 - протостель; 2 - сифоностель; 3 - сифоностель с расчлененной ксилемой; 4 - эустель.
или соленостелью1 (рис. 182), в отличие от эктофлойной сифоностели (рис. 182), имеющей флоэму только кнаружи от ксилемы. Параллельно с образованием ответвлений стели в боковые ветви стебля и в листья происходит переход конструкции стели от цилиндрической или полуцилиндрической к сетчато-полуцилиндрической. Эволюция стели в этом направлении произошла у представителей большой группы высших растений: папоротникообразных (Pteropsida), голосеменных (Gymnospermae) и покрытосеменных (Angiospermae) - и характеризуется тем, что проводящая система ветви и листа отражается на строении стели несущего их органа, вызывая образование в стели прорывов, или брешей. Вместе с тем усиливается и витализация стели: под эндодермой дифференцируется паренхиматический перицикл. Во флоэме и ксилеме появляется паренхима. Каждая ситовидная трубка и каждый сосуд входят в непосредственное соприкосновение с живыми клетками. В области листовых прорывов внешняя и внутренняя эндодермы смыкаются как в стели стебля, так и в отходящем листовом следе; сердцевина стебля входит в прямое сообщение с его первичной корой и с сердцевиной листового черешка.
На сильно облиственных стеблях, т. е. при тесном расположении листовых прорывов, соленостель представляет трехмерную сетку, имеющую на поперечных разрезах вид кольца из перерезанных звеньев стели (меристелей2) с участками основной ткани в промежутках. Такая соленостель называется диктиостелью3 (рис. 181, В; 182, 5; 183). На более высокой стадии морфогенеза диктиостели в каждый лист отходит (например, у папоротника мужского) по нескольку тяжей, прикрепляющихся к нижнему и боковому краям листового прорыва.
Дальнейшие этапы эволюции стели - образование перфорированной и полициклической диктиостели. У более высокостоящих папоротников в меристелях диктиостели образуются
211
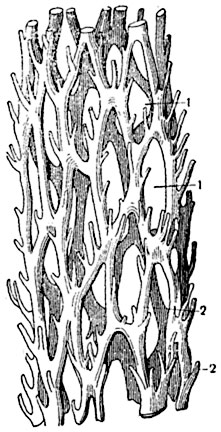
Рис. 183. Отпрепарированный отрезок диктиостели мужского папоротника (Dryopteris filixmas), в перспективном изображении:
1 - листовые бреши; 2 - обрезанные листовые следы.
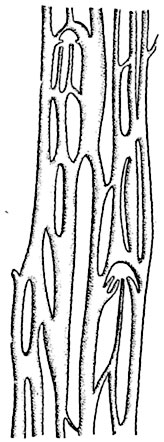
Рис. 184. Отпрепарированный и развернутый в одну плоскость отрезок диктиостели с перфорациями (с "окнами") папоротника Stenochlaena tenuifolia.
перфорации - прогалы, обычно удлиненно-овального очертания, независимые от листьев и их следов и соединяющие сердцевину с корой. Края перфораций покрыты, как и в листовых прорывах, эндодермой, смыкающейся с внешней и с внутренней эндодермой стели. Перфорированная стель имеет в целом вид частой сетки, а на поперечных разрезах представляется раздробленной на многочисленные мелкие тяжи. Такого рода стель образуется преимущественно в крупных длинных горизонтальных корневищах (у видов рода Platycerium), реже - в отвесных стеблях (Stenochlaena tenuifolia, рис. 184).
При полициклии внутри диктиостели образуются добавочные, стелярные структуры в виде полых цилиндров с прорывами: внутренний цикл иногда представлен всего лишь одним тяжом (рис. 185). Стелярная система и при полицикличности является единой стелью: ее конструктивное единство может быть обнаружено в базальных частях органа, где та же стель не расчленена на циклы. Полицикличность стели связана с размерами и - косвенно - с возрастом органа: так, например, у папоротника Matonia pectinata корневище особи постепенно, с возрастом, образует один, затем два, три добавочных стелярных цилиндра (цикла). У более мелкого вида Маtonia (M. sarmentosa) образуется лишь один полный и затем второй, неполный добавочный цикл.
У крупного ископаемого папоротника Psaronium infractus в стелярной системе с поперечником в 64 мм было насчитано не менее 12 циклов. Внутренние циклы имеют листовые прорывы, аналогичные прорывам наружной стелярной сетки. Строение полициклических стелей может усложняться образованием анастомоз - перемычек между циклами. Усложнение стели наблюдается в онтогенезе (рис. 186).
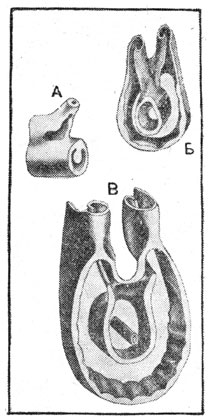
Рис. 185. Перспективные изображения отрезков полициклической стелярной системы папоротника Matonia pectinata:
А - стель молодого стебля (в узле); Б - стель более взрослого стебля (вид на узел сзади); В - стель еще более взрослого стебля, вид на узел почти в плане (рисунки с восковых моделей).
212
У плауновидных (Lycopsida) и клинолистных (Sphenopsida), которые сохранились в наше время в виде сравнительно мелких травянистых представителей - плаунов, селагинелл и хвощей, листья не влияют на архитектонику стели стебля. У всех у них в листья отходят проводящие тяжи, но эти тяжи только примыкают к выступам на периферии стели, не вызывая в ней образования прорывов. Это относится даже к таким Lycopsida, которые, как вымершие сигиллярии, имели листья крупные, длиной свыше 1 м. Морфологический к физиологический прогресс в филоморфогенезе стели у Lycopsida в принципе тот же, что и у Pteropsida: развитие стели шло по пути витализации - увеличения поверхности соприкосновения между проводящими тканями и живыми клетками.
У плауновых выработались, однако, структуры стели с ксилемой, составляющей не сетку, а губку. Чтобы представить себе структуру этого рода, надо рассмотреть поперечные сечения стели различных видов плауна (рис. 187) и принять к тому же во внимание, что тяжи ксилемы не идут вдоль стебля параллельно его продольной оси, а изгибаются и соединяются друг с другом. То же можно сказать о флоэме и паренхиме. Стель с системой ксилемных тяжей в виде губки называют плектостелью4. Некоторые варианты плектостели на поперечных разрезах имеют сходство с актиностелью (рис. 187, А, Б). У селагинелл стель эволюционировала из протостели: а) в стель, разветвляющуюся на несколько продольных меристелей с поперечным сечением в виде широких, изогнутых и лопастных полосок (у Selaginella wildenovii) или небольших кругов или овалов, или же б) в стель полициклическую, состоящую на поперечных разрезах из двух отдельных, приблизительно концентрических стелярных кругов (у S. lyallii, рис. 188).
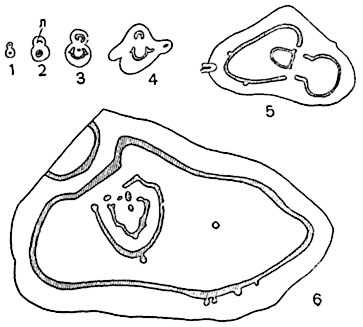
Рис. 186. Усложнение конфигурации стели в ходе онтоморфогенеза, в коническом стебле папоротника (Pteris polyphylla).
Серия поперечных разрезов стебля на различных расстояниях от его основания; все разрезы изображены в одном масштабе, стель заштрихована; 1, 2 - протостель; л - листовой след; 3, 4 - форма стели более сложна; 5, 6 - стель полициклична.
В индивидуальном, онтогенетическом развитии организмы повторяют отдельные этапы их филогенетического развития, в том числе и отдельные этапы образования той или иной сложной стели из протостели. В работах профессора МГУ К. И. Мейера и его учеников прослежено постепенное образование стели папоротников и хвощей; исследовано развитие проводящей системы проростков, вырастающих непосредственно из оплодотворенной яйцеклетки - зиготы, и постепенное формирование стели из меристемы конуса нарастания. Как у папоротников, так и у хвощей развитие начинается с протостели, затем на очень коротком участке (около 1 мм) происходит переход к сифоностели и затем диктиостели. Молодой проросток имеет протостелическое строение (рис. 189).
Усложнение стели в одном и том же стебле в направлении от его основания к вершине можно заметить на серии поперечных разрезов стебля на разных его уровнях.
213
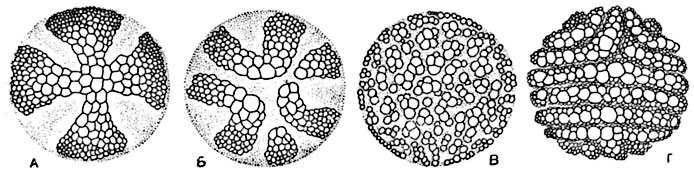
Рис. 187. Стели четырех видов плауна (Lycopodium) на поперечных разрезах (полусхематично).
В ксилеме изображены поперечные сечения трахеид, флоэма отмечена точками. А - стель Lycopodium serratum, с очертанием ксилемы в виде креста; Б - стель прямостоячего стебля L. annotinum, с лопастно-звездчатой конфигурацией ксилемы; В - стель L. cernuum, с мелкими ксилемными группами, разбросанными равномерно по всему сечению стели; Г - стель L. volubile, с билатеральным строением.
Стель покрытосеменных. Переходя к стеблям двудольных и однодольных, мы можем сказать, что их первичные тканевые образования располагаются по типу диктиостели. Происхождение стели стеблей двудольных с пучковым строением можно представить так. В стели, находящейся на ступени эктофлойной сифоностели (рис. 182), ксилемная трубка витализируется как бы внедряющимися в нее со стороны сердцевины продольными прослойками паренхимы и расчленяется на продольные ксилемные тяжи (рис. 182); флоэма же образует по-прежнему сплошную трубку; получается стель того типа, как у папоротника Osmunda. У последнего от листовых прорывов в стели отходит лишь по одному листовому следу, направляющемуся в лист от нижней стороны прорыва. В случае образования во флоэмной трубке продольных прослоек основной паренхимы, составляющих вместе с прослойками в ксилеме первичные сердцевинные лучи, получается хорошо известный тип стели - эустель5 (рис. 182).
У большинства однодольных и у некоторых двудольных стель формируется в виде атактостели (рис. 131), для которой характерно большое число меристелей в виде проводящих пучков, сильно изогнутых на большей части длины и объединяющихся в сетку путем слияния их ответвлений в узлах и в междоузлиях; на поперечных разрезах междоузлий стебля проводящие пучки представляются разбросанными по всей площади сечения - отсюда термин "атактостель"6 для стели однодольных. При эустелии и при атактостелии эндодерма может окружать всю стель в целим или гораздо реже - проводящие пучки. Многими однодольными эндодерма утрачена.
У голосеменных и у большинства двудольных стель дополняется и усложняется вторичным приростом - образованием вторичных элементов ксилемы и флоэмы. В расположении во вторичном приросте одревесневших и неодревесневших элементов, межклетников и т. д. получается конфигурация сетки, имеющая выгоды, о которых упоминалось.
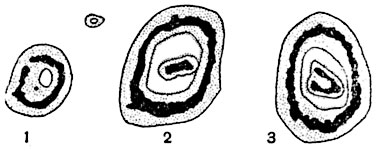
Рис. 188. Серия последовательных поперечных срезов стели корневища Selaginella lyallii (от основания к верху):
1 - соленоксилия; 2 - соленостелия; 3 - полициклия.
На всех фигурах ксилема зачернена.
214
У наиболее прогрессивных покрытосеменных - у травянистых растений - витализация стели чрезвычайно сильна не только в первичных тканях, но и у трав с вторичным приростом и во вторичных тканевых образованиях: в древесине и в лубе образуется в большом количестве живая паренхима - продольная и (в древесинных и лубяных лучах) поперечная.
Сетчатость и рубчатость стели имеют еще значение в облегчении газового обмена стелярной паренхимы с корой и далее через пнейматоды (устьица, чечевички) с наружной атмосферой. Раздробленность проводящих и арматурных тканей при сетчатой (и губчатой) структуре стели и удержание вместе с тем связанности и сращения их с другими тканями являются важным приспособлением и с точки зрения современного учения о строительной механике растения.
Например, прямостоячий побег Selaginella lyallii, имеющий в верхней части резко выраженную диктиостель, в эксперименте принуждали продолжать рост и развитие в горизонтальном положении. Оказалось, что стель в части побега, образовавшейся при измененных условиях, меняла постепенно характер конструкции и возвратилась в итоге к соленостелическому строению.
Ветви и листья сильно влияют на форму стели. Рассматривая сосудистые пучки, соединяющие стебель с его ветвями и листьями, мы встречаемся со следами ветвления, прорывами ветвления, листовыми следами и листовыми прорывами. Наиболее ясно их влияние бывает выражено при трубчатом строении стели, когда флоэма окружает трубчатую ксилему (сифоностелия) или в некоторых случаях эустелии.
Прорывы ветвления и следы ветвления. Из ствола в ветви и вообще в оси (n + 1)-го порядка из осей n-го порядка отходят меристели в количестве одной, реже двух. Эти стелярные ответвления, обычно содержащие по одному крупному проводящему пучку, называются следами ветвления (рис. 190) или рамулярными7 следами. Если в ответвление идет одна меристель, она имеет сечение в форме подковы или полулуния, обращенного выпуклой стороной кверху. Недалеко от места выхода из стели в ответвления меристель становится замкнутой, цилиндрической.
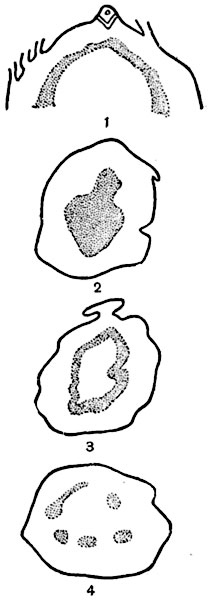
Рис. 189. Усложнение конфигурации стели в онтогенезе стебля папоротника (Polypodium postulatum).
Продольный срез (1) и поперечные срезы стебля на различных расстояниях от точки роста; все срезы изображены в одном масштабе, меристема проводящей системы обозначена точками; 2 - протостелия; 3 - соленоcтелия; 4 - диктиостелия.
Замкнутая стель образуется в ветви и при двух рамулярных следах путем их слияния и увеличения их размеров по окружности поперечного сечения. Рамулярные следы обычно резко по короткой кривой отходят от стели оси.
215
Прорывы ветвления вызывают в стели ствола значительные и длительные местные изменения8. Сердцевина ветви остается соединенной с сердцевиной ствола. Древесинное тело ветви приобретает форму конуса с вершиной, обращенной в сторону сердцевины ствола. Годичные кольца древесины ветви соединяются с годичными кольцами древесины ствола. При этом на верхней стороне той части древесинного тела ветви, которая находится внутри ствола, в месте смыкания древесины ствола и ветви образуются складки. По мере роста ствола в толщину длина захороненной части древесинного тела ветви увеличивается. Луб, будучи зажат с двух сторон и подвергнут давлению с двух противоположных сторон, под напором нового, вторичного прироста выжимается из пазухи, Он испытывает не только радиальное растягивающее его напряжение, как луб свободной части любой оси, но и напряжение, выталкивающее его из пазухи. В результате луб сильно деформируется: старый луб получает трещины, молодой ложится в складки. В древесине в месте соединения ветви со стволом обычно складок и трещин не образуется, благодаря тому что здесь древесина формируется из более коротких гистологических элементов и что при развитии волокон может иметь место скользящий рост. У некоторых пород, как например у дубов, все же близ основания энергично утолщающихся ответвлений образуется волнистая древесина.
Прорывы ветвления довольно крупны и простираются на значительное расстояние вдоль органа. Нередко прорыв ветвления объединяется с листовым прорывом.
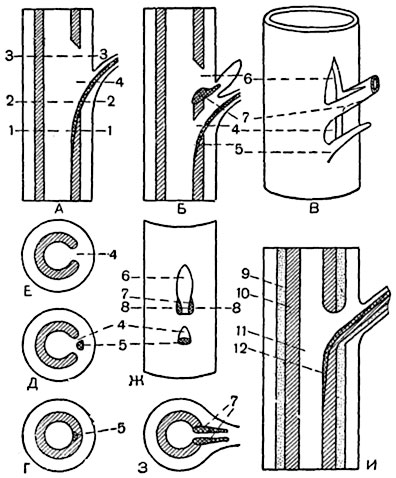
Рис. 190. Схемы прорывов ветвления и листовых следов:
А - продольный разрез через стебель с листовым следом (2) и листовым прорывом (4); Б - продольный разрез; В - перспективное изображение отрезка стебля с листовым следом (5) и листовым прорывом (4), со следом ветвления (7) и прорывом ветвления (6). На фиг. Б - боковая ветвь в стадии почки; на фиг. В - в развитом состоянии; Г, Д, Ε - поперечные сечения стебля, изображенного на фиг. А , на уровнях 1-1, 2-2, 3-3; Ж - вид наружной поверхности участка стебля В после отрезания у поверхности следов листа и ветви; З - поперечный разрез через отрезок стебля (В) на уровне 8-8, показанном на фиг. Ж. На фиг. А-З не показаны раздельно флоэма и ксилема; И - более детальная схема продольного разреза через стебель с листовыми следом и прорывом: 9 - луб, 10 - древесина, 11 - сердцевина, 12 - ксилема следа.
Листовые следы и листовые прорывы. У голосеменных и у покрытосеменных из стебля отходит в каждый лист одна или несколько меристелей, включающих по одному или несколько проводящих пучков и называемых
216
листовыми следами9. Число листовых следов обычно является постоянным для данного вида или для более крупной систематической единицы. Число это вместе с тем не находится в прямой зависимости от величины листа и от ширины его основания: например, крупные листья ясеней имеют один, а мелкие кроющие листья в соцветиях у ив - три листовых следа. Листовой след включает, как правило, тяж протоксилемы и тяж протофлоэмы; кроме того, в нем могут иметься метаксилема, метафлоэма и вторичные проводящие ткани. За верхнюю границу листового следа принято считать участок его в основании черешка листа. На нижнем конце листовой след сливается с другим листовым следом или со стелью стебля.
Листовой след бывает почти прямым и входящим радиально по отношению к стеблю (рис. 191, Н) или же изогнутым (рис. 191, О-С); нередко изогнутый листовой след обладает двоякой кривизной: таковы, например, опоясывающие листовые следы (рис. 191, И-М), отходящие из стебля в листья, прикрепляющиеся широким основанием.
Листовые следы нередко разветвляются или сливаются в области первичной коры стебля. Здесь, например, обычно ответвляются от боковых листовых следов проводящие пучки в прилистники. В листе листовые следы продолжаются в проводящие пучки, разветвляющиеся и сливающиеся друг с другом. В листовом черешке преобладает слияние, в пластинке - разветвление пучков. Листовой след обособляется пространственно от проводящей системы стебля обычно лишь постепенно, и некоторые листовые следы тянутся по стеблю на значительной длине, не выходя из границ флоэмноксилемного остова стели. При своем основании листовой след дифференцируется как сектор, состоящий преимущественно из протоксилемы и протофлоэмы, различимый под микроскопом (рис. 191, А, Г); выше он обособляется с двух сторон (рис. 191, В), а еще выше выходит в направлении к листу (рис. 191, Б). В положении, показанном на рисунке 191, А, листовой след может проходить через одно или несколько междоузлий.
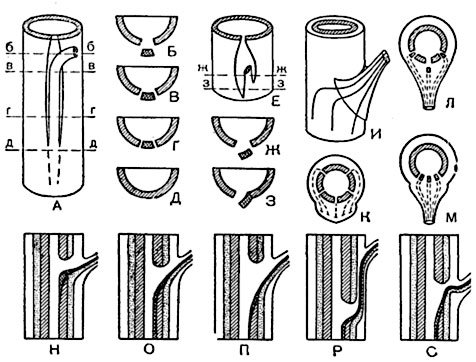
Рис. 191. Схемы пространственных отношений листовых следов и стели у некоторых двудольных и голосеменных.
А-Д - листовой след, обособляющийся от стели ниже места выхода следа в лист: А - общий вид в перспективе; Б, В, Г, Д - поперечные разрезы на уровнях б-б, в-в, г-г, д-д; Е, Ж, З, как в А-Д, но след обособляется с одной стороны ниже, чем с другой: Е - общий вид, Ж и З - поперечные разрезы на уровнях ж-ж и з-з; И, К - опоясывающие листовые следы: И - в перспективе (с указанием пунктов выхода следов из цилиндра стели стебля и мест вступления их в листовой черешок), К - поперечное сечение стебля со стелью и листовые следы (до выхода в черешок), в проекции на плоскость сечения; Л и Μ - схемы, аналогичные К: Л - для случая трех листовых следов, отходящих от стели стебля раздельно; Μ - для случая трех следов с общей листовой брешью; Η-С - продольные разрезы стеблевых узлов, с различной кривизной листовых следов; ксилема отмечена косой штриховкой, флоэма - точками.
217
Листовой след у основания его обычно симметричен, но в некоторых случаях отщепляется с одной стороны ниже, чем с другой, и тогда кажется отходящим не от основания прорыва в проводящем остове стели, а от одного из боков прорыва (рис. 191, Ε-З).
В отличие от папоротникообразных листовые прорывы голосеменных и покрытосеменных сравнительно малы: стель оси смыкается уже на небольшом расстоянии от места отхождения листового следа. Если лист имеет несколько листовых следов, то они отходят обычно раздельно (рис. 191, И-Л), реже - друг подле друга; в последнем случае им соответствует один общий прорыв в стели (рис. 191, М).
218
1 "Сифон", "солен" по-гречески - трубка. Некоторые авторы считают термины "сифоностель" и "соленостель" синонимами, другие называют соленостелью только амфифлойную сифоностель.
2 От греческого "мерис" - доля, часть.
3 От греческого "диктион" - сетка.
4 От греческого "плектос" - сплетенный.
5 От греческого "эу" - хороший, настоящий.
6 От греческого "атактос" - беспорядочный.
7 По-латыни ramulus - ветка.
8 Структуры в месте соединения ответвления любого (n + 1)-го порядка с несущей его осью n-го порядка в основных чертах сходны. Ради краткости будем говорить только о соединении оси 2-го порядка с несущей его осью 1-го порядка.
9 Нередко листовым следом называют весь комплекс пучков, продолжающихся из стебля в лист.