

ПЛАСТИДЫ
Пластиды всегда находятся в протоплазме, близки к ней по физическим и химическим свойствам, возникают только от пластид. Они способны к росту и размножаются делением, могут образовывать в своем теле (в строме) определенные пигменты и формировать внутри стромы крахмал. В зависимости от содержания тех или иных пигментов находится окраска и функции основных пластид высших растений: а) зеленых пластид (хлоропластов), б) красных и желтых (хромопластов) и в) бесцветных (лейкопластов).
Все пластиды, по-видимому, имеют сходное строение; лучше изучено строение хлоропластов.
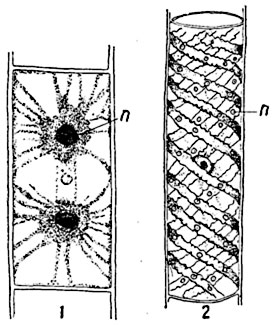
Рис. 28. Формы хлоропластов водорослей:
1 - клетка из нити Zygnema cruciatum; 2 - отдельная клетка из нити спирогиры; п - пиреноиды.
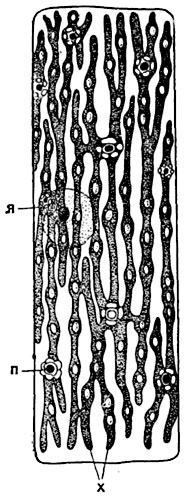
Рис. 29. Сетчатый хроматофор в клетке водоросли эдогониум:
х - хроматофор; п - пиреноиды с крахмалом; я - ядро.
Хлоропласты. Хлоропласты высших растений (называемые также хлорофилловыми зернами) по форме до некоторой степени сходны с линзами: в плане хлоропласт имеет очертание, более или менее близкое к кругу, а при рассматривании в профиль напоминает эллипс. Если хлорофилловые зерна лежат тесным слоем, то, нажимая друг на друга, они принимают угловатую форму. Число хлорофилловых зерен в различных клетках очень изменчиво. Например, в клетках листа клещевины количество хлорофилловых зерен колеблется от 10 до 36, в клетках Elodea densa - от 26 до 32. Диаметр хлорофилловых зерен составляет 4-9 μ.
По своим размерам хлорофилловые зерна менее разнообразны, чем ядра, а тем более сами клетки, хотя некоторые авторы отмечают, что хлорофилловые зерна крупнее в клетках больших размеров. Когда И. И. Герасимов в культурах спирогиры получал крупные клетки с двойной массой ядра, то в таких клетках и спиральные хлоропласты были крупнее, чем в нормальных, и число их возрастало с 8 до 12-13 (рис. 18). Исключительно крупные хлоропласты отмечены для Peperomia metallica: диаметр
58
хлоропластов достигает 24 μ, но здесь следует отметить, что число их в клетке очень невелико - их только 4.
Хлоропласты могут изменять форму и размеры. Некоторые изменения зависят от поверхностного натяжения; при возрастании его уменьшается величина поверхности, и форма пластиды приближается к сферической - пластида "округляется"; при уменьшении поверхностного натяжения пластида удлиняется. Форма хлоропластов может меняться в зависимости от освещения; например, в листьях клещевины, подвергнутых затенению, хлоропласты становятся почти изодиаметрическими (с наибольшим размером ∼6,3 μ и наименьшим ∼5,7 μ); на ярком свету они меняют форму на чечевицеобразную (с диаметром ∼8,3 μ и толщиной ∼3,6 μ).
В культуре водяной чумы (элодеи) при разных температурах хлорофилловые зерна в листьях, выросших при более высокой температуре, получались почти вдвое меньшими.
Очень разнообразны по форме хлоропласты водорослей - хроматофоры (рис. 28, 29).
В окрашенных пластидах - хроматофорах - многих водорослей (рис. 28, 29) и некоторых из печеночников, относящихся к роду Anthoceros, имеются особые, тягуче-жидкие тельца, чаще всего округлой или угловатой формы; эти тельца, называемые пиреноидами, богаты белковыми веществами, но нуклеинов не содержат. Вокруг пиреноидов обычно располагаются мелкие крахмальные зерна; эти зерна крахмала образуются в клетке в первую очередь, а исчезают в последнюю. Пиреноиды образуются путем деления уже существующих, но могут и возникать в клетке заново.
Хлоропласты содержат в строме четыре пигмента: два зеленых1 (хлорофилл a и хлорофилл b), оранжево-красный (каротин, или, иначе, каротен) и желтый (ксантофилл).
По своему химическому составу хлорофилл представляет собой сложный эфир дикарбоновой кислоты хлорофиллина и двух спиртов - метилового и фитола.
Хлорофилл а: |
C32H30ON4Mg |
╱ ╲ |
COOCH3 COOC20H39 |
Хлорофилл b: |
C32H28O2N4Mg |
╱ ╲ |
COOCH3 COOC20H39 |
Хлорофилл а отличается от хлорофилла b по количеству атомов водорода и кислорода.
Хлорофилл а имеет синеватый оттенок, хлорофилл b - желтоватый. Молекулярный вес хлорофилла равен ∼ 900.
И. П. Бородин, обрабатывая срезы зеленых частей растения на предметном стекле этиловым спиртом, получал после медленного высушивания препарата темно-зеленые или почти черные кристаллы в виде трех- или шестиугольных пластинок и тетраэдров. В дальнейшем было выяснено, что это кристаллы хлорофилла, в молекулах которого фитольная группа замещена этильной.
Центральное место в молекулах хлорофиллов a и b занимает атом магния, связанный с 4 атомами азота.
Содержание хлорофилла в нормальных зеленых листьях у самых разнообразных растений составляет примерно 0,8 % сухого веса (0,3-0,7 г на 1 м2 листовой поверхности).
59
В одном хлорофилловом зерне содержится 6% хлорофилла; остальное составляют вода, белки, липиды и др.
Из пигментов, сопровождающих хлорофиллы, оранжево-красный каротин представляет ненасыщенный углеводород формулы C40H56, а желтый ксантофилл (C40H56O2) - двухатомный спирт, как бы продукт окисления каротина. Каротин и ксантофилл относятся к обширной группе каротиноидов - пигментов желтого, оранжевого и красного цветов, ряд других представителей которых также встречается в растениях. Всю совокупность пигментов хлоропласта иногда называют "хлорофиллом" в широком смысле слова.
Вытяжка зеленых листьев и в меньшей мере сами листья обнаруживают явление флуоресценции. В проходящем свете вытяжка кажется зеленой, а в падающем - вишнево-красной. За очень редкими исключениями, в органах покрытосеменных растений при развитии их в отсутствии света зеленые пигменты в хлоропластах не образуются или образуются в ничтожном количестве; на свету происходит их быстрое позеленение.
В хлоропластах совершается сложный процесс фотосинтеза - образования углеводов из углекислого газа и воды под действием энергии солнечного света2.
Окончательный результат процесса фотосинтеза можно представить в виде следующей реакции:
6CO2 + 6H2O + 674 ккал → C6H12O6 + 6O2
Образовавшийся углевод обычно полимеризуется в крахмал по схеме:
nC6H12O6 → (C6H10O5) n + n H2O
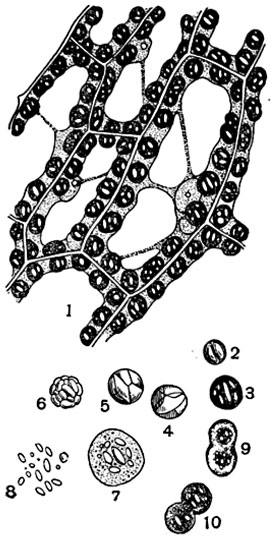
Рис. 30. Хлорофилловые зерна листа мха Funaria hydrometrica:
1 - участок взрослого листа с несколькими клетками (в плане); в постенном слое протоплазмы расположены хлорофилловые зерна с мелкими крахмальными зернами (отмечены белым цветом); 2-10 - отдельные хлорофилловые зерна с крахмалом: 2 - молодое, 3 - более взрослое, 9 и 10 - делящиеся, 4, 5 и 6 - заполненные крахмалом, 7 - молодое, набухшее в воде, 8 - расплывшееся в воде и оставившее после себя крахмальные зерна.
Крахмал откладывается в хлоропластах в виде мелких зерен ассимиляционного, или автохтонного, крахмала (рис. 30, 32).
У некоторых растений, преимущественно однодольных, ассимиляционного крахмала обычно не образуется (кроме как в замыкающих клетках устьиц) и продуктом фотосинтеза является глюкоза. При сильно повышенном содержании CO2 в атмосфере в хлоропластах сахарообразующих растений (сахарного тростника, сахарной свеклы) на свету появляется крахмал.
Способность к фотосинтезу и ряд других свойств пластид как биологически активных систем объясняются наличием в хлоропластах ферментов. Эта очень сложная ферментативная система обеспечивает не только
60

Рис. 31. Хлоропласт из клеток кукурузы (электронный микроскоп, увел. 40 000).
весь процесс фотосинтеза, но и отток продуктов ассимиляция из хлоропласта. В состав этой системы входит зеленый пигмент пластид - хлорофилл.
О внутренней структуре хлоропласта было очень много споров, выдвигалось много теорий, но только применение электронного микроскопа дало возможность более детально изучить его субмикроскопическое строение (рис. 31, 32). В настоящее время считают, что хлоропласты высших растений имеют пластинчатую структуру. Пластинки стромы чередуются с пластинками, состоящими из гранул (зернышек), содержащих хлорофилл. Связь между гранулами и стромой в настоящее время еще не совсем ясна.
В выяснении вопросов роли хлорофилла в процессе фотосинтеза и значении лучей различных областей солнечного спектра в этом процессе большая заслуга принадлежит К. А. Тимирязеву. Тимирязев изучал хлорофилл как "связующее звено между солнцем и жизнью", а хлорофилловое зерно - как тот фокус, ту точку в мировом пространстве, где солнечный луч, превращаясь в химическую энергию, становится источником всей жизни на Земле.
Тимирязев установил, что наиболее интенсивно поглощаются красные лучи (с длиной волны от
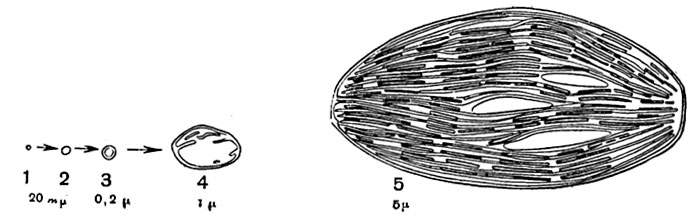
Рис. 32. Схематическое изображение развития хлоропласта.
Справа - сформированный хлоропласт, в строме которого видны три линзовидных крахмальных зерна.
61
730 до 680 мμ) и в несколько меньшей мере лучи сине-фиолетовой части спектра (с длиной волны 470 мμ и меньше).
Осенью перед опадением листа хлорофилловые зерна желтеют; это пожелтение зависит от того, что зеленые пигменты разрушаются раньше, чем желтые; пожелтение связано с оттоком веществ из листовой пластинки в осевые органы.
Хромопласты. Хромопласты - пластиды, содержащие пигменты из числа каротиноидов (каротин и ксантофилл). Они имеют окраску от желтой (в лепестках лютиков) и оранжевой (в кожуре апельсинов) до оранжево-красной (в корнях моркови) и ярко-красной (в плодах шиповников).
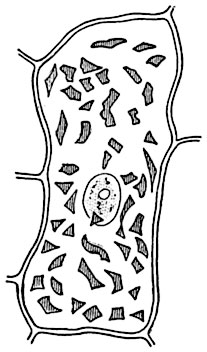
Рис. 33. Клетка чашелистика настурции Tropaeolum majus с ядром и хромопластами.
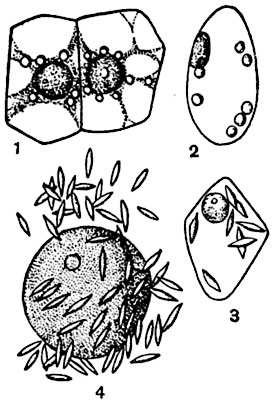
Рис. 34. Лейкопласты:
1 - в клетках кожицы Philodendron grandifolium; 2, 3 - в клетках семени Melandrium macrocarpum; 4 - в клетках корня Phajus grandifolius (изображено лишь ядро с лейкопластами подле него).
Хромопласты встречаются преимущественно в клетках частей цветка, чаще всего в лепестках (у лютиков, настурций) или в листочках венчиковидного околоцветника (у некоторых лилий, касатиков); имеются они и в плодах многих растений (шиповников, рябин, ландышей). Окраска хромопластов делает цветки и плоды заметными издали, т. е. способствует опылению растений и распространению плодов и семян птицами и млекопитающими. Сравнительно небольшое число растений содержит хромопласты в клетках вегетативных органов. В качестве примеров приведем мясистые корни и гипокотили моркови, надземные органы некоторых паразитов (заразихи, гнездовки).
По форме хромопласты весьма разнообразны. Изредка они бывают эллипсоидальными или при тесном расположении многоугольно-таблитчатыми, лопастными и т. д. Обычно хромопласт имеет игловидное и угловатое очертание, его строма растянута пигментом, составляющим преобладающую по объему часть хромопласта (рис. 33). У многих растений в хромопластах отмечается наличие крахмала.
62
В хромопластах иногда обособляются белковые вещества или образуются капли масла: в клетках кожуры плодов апельсина и других цитрусовых пигмент хромопластов частично растворен в эфирных маслах.
Не всегда окраска плодов зависит только от хромопластов. От наличия пластид зависят цвета зеленый, желтый, кирпично-красный (лепестки цветков лютиковых, сложноцветных, корни моркови, многие созревающие плоды). Цвета же синий, малиновый, темно-красный (плоды малины, калины) зависят от окраски клеточного сока, содержащего антоцианы. Часто получается смешанный цвет, обусловленный окраской клеточного сока и пластид.
Очень показательно провести отделение каротина бензолом от спиртовых вытяжек плодов красного перца и рябины. Если к спиртовой вытяжке плодов перца добавить бензол, то сверху в бензоле окажется растворенным каротин, а ксантофилл окажется внизу в спирте. Клеточный сок (также остается в спирте) бесцветный. Если повторить этот опыт с плодами рябины, можно видеть, что каротина в них не так уж много, так как бензольная фракция будет гораздо более бледной, чем таковая красного перца, а спирт останется окрашенным в розовый цвет от наличия антоциана.
Белый цвет лепестков обусловлен равномерным рассеиванием света при отражении его от пузырьков воздуха, заключенных в сильно развитых межклеточных пространствах и на поверхности органа.
Желтый цвет некоторых цветков и плодов (георгин, льнянка, мак, лимон) зависит также от пигмента клеточного сока, родственного антоциану, - антохлора.
Окраска частей растений может зависеть еще от цвета отмерших клеток и их измененного содержимого (оболочка семян, поверхность стволов деревьев).
Лейкопласты. Пластиды, не содержащие в строме пигментов и называемые лейкопластами, имеются во многих клетках большинства растений (рис. 34). Так как лейкопласты бесцветны и к тому же преломляют свет почти так же, как протоплазма, не всегда легко обнаружить их присутствие в клетке.
По форме лейкопласты обычно почти шаровидны. В тех случаях, когда в их строме находится крахмал или белок, они принимают иные очертания. Сравнительно богаты лейкопластами образовательные ткани, подземные органы, семена.
В лейкопластах может образовываться крахмал, отлагающийся в виде зерен в их строме.
Во многих случаях крахмал накопляется в лейкопласте в столь большом количестве, что живое тело пластиды (строма) оттесняется на периферию. Его можно не без труда заметить в виде очень тонкой пленки на поверхности крахмального зерна; в этих случаях лейкопласт является крахмалонакопителем в полной мере (подробнее о запасном крахмале см. на стр. 78).
Движение пластид. Кроме перемещений пластид, связанных с распределением их между дочерними клетками, образующимися в результате деления клетки, совершаются другие передвижения их: 1) зависящие от возрастных изменений клетки и пластид и 2) обратимые и многократно повторяющиеся движения, связанные с изменениями в направлении и интенсивности воздействия факторов среды ("ориентировочные" движения пластид).
Хлорофилловые зерна, находящиеся в постенном слое протоплазмы, могут перемещаться в клетке. Они не только пассивно увлекаются протоплазмой при ее движении, но, в зависимости от силы и направления света, сами могут менять свое положение в клетке. И. П. Бородин показал, что лучше всего это видно на тонких листочках ряски (Lemna trisulca).
На рисунке 35 изображено размещение хлорофилловых зерен в клетках, которые находились в различных условиях освещения. На рассеянном свету хлорофилловые зерна располагаются по стенкам, которые находятся под прямым углом к главному направлению лучей; на каждое зерно непосредственно попадает бо́льшая часть падающего света. На ярком солнечном свету хлорофилловые зерна перемещаются на боковые стенки, которые лежат
63
вдоль падающих лучей. Свет, проникающий в клетку, не попадает на зерно прямо, а рассеивается в стороны и освещает при этом зерна более равномерно. В темноте зерна располагаются по стенкам, которые соприкасаются со стенками соседних клеток, как говорят по теплым стенкам, но последнее расположение не всегда удается наблюдать.
Интересно, что освещение отдельных хлорофилловых зерен происходит весьма равномерно. Самостоятельная скорость перемещения хлорофилловых зерен в клетке равна 0,12 μ в секунду, т. е. скорость значительно меньшая, чем движение амебы и плазмодия.
На расположение хлорофилловых зерен влияет не только сила и направление света. В некоторых случаях наблюдалось перемещение хлорофилловых зерен к тем участкам клеточной стенки, к которым подходят межклеточные пространства; по межклетникам поступает углекислота, поглощаемая в процессе ассимиляции.
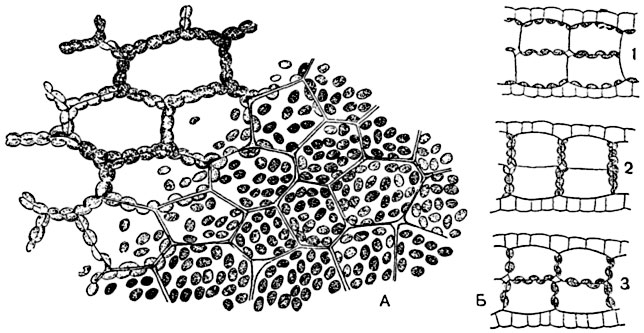
Рис. 35. Перемещение хлорофилловых зерен в клетках ряски трехдольной (Lemna trisulca):
А - часть растения в плане, верхняя левая часть находилась в темноте, правая нижняя часть - на рассеянном свету (по Бородину); Б - частичные поперечные разрезы через растения, выдержанные: 1 - на рассеянном свету; 2 - на прямом солнечном; 3 - в темноте.
Происхождение пластид. Резких границ между пластидами различных категорий нет. Пластиды одного типа могут переходить в пластиды другого типа. В самом деле, в проростках и во взрослых особях многие из пластид, возникающих в результате деления лейкопластов клеток зародыша, становятся хлоро- и хромопластами. Хлоропласт, теряя хлорофилл и обогащаясь каротиноидами (как в созревающих плодах шиповника), становится хромопластом; лейкопласт при накоплении в его строме каротиноидов превращается в хромопласт (как в корнях моркови). Достаточно обнажить верхнюю часть растущего в почве красного "корнеплода" моркови от земли, чтобы через некоторое время эта часть гипокотиля и корня позеленела: хромопласты клеток становятся хлоропластами.
Вопрос о первоначальном возникновении пластид в клетках растений в настоящее время еще не совсем ясен. Существовала теория возникновения пластид из хондриосом; большинство исследователей от нее отказались. В последнее время высказывается мнение, что пластиды возникают из особых образований - пропластид.
64
1 В настоящее время известно, что в растениях хлорофилл присутствует в нескольких формах, но известно строение только двух его форм - a и b, причем и здесь положение двойных связей и характер связи магния с азотом еще нуждаются в уточнении.
2 А. С. Фаминцын впервые установил, что процесс фотосинтеза может происходить и при искусственном освещении.