

Строение типичного зеленого листа
В первую очередь мы рассмотрим строение типичных зеленых листьев, имея в виду главным образом листья мезофитов, т. е. растений, приуроченных к местообитаниям со средними условиями увлажнения, например обитающих на лугах. В связи с выполнением функций фотосинтеза и транспирации, зеленый лист целиком, если он сидячий, или в значительной мере, если он черешковый, представляет собой тонкую пластинку с поверхностью, значительной по отношению к объему.
В пластинке листа уже с помощью лупы можно различить 4 группы тканей: 1) покровную - кожицу, или эпидермис; 2) основную, питательную - мезофилл1; 3) проводящую - сосудисто-волокнистые пучки (жилки); 4) механическую, придающую листу жесткость, определяющую положение листа в пространстве.
Эпидермис. Лист окружен со всех сторон покровной тканью - эпидермисом, или кожицей (рис. 174). Со строением кожицы вы уже ознакомились раньше, при изучении тканей (стр. 109).
Эпидермис стебля переходит на черешок и пластинку листа. Кожица состоит из одного слоя таблитчатых, плотно сомкнутых клеток без межклетников. Наружные стенки этих клеток сильно утолщены и покрыты кутикулой. Клетки кожицы живые; они не содержат хлорофилловых зерен. Местами, преимущественно на нижней стороне листа, в эпидермисе находятся устьица.
На поверхности листа клетки кожицы не везде одинаковы: клетки кожицы, покрывающей нижнюю сторону листа, обычно более мелки и их стенки более извилисты, чему клеток верхней кожицы (рис. 86). Извилистость очертаний клеток и верхней и нижней кожицы усиливается при повышении влажности воздуха, в котором развивается лист. Верхняя кожица отличается от нижней еще и большей кривизной наружных стенок клеток и большей их толщиной. Мощность слоя воска, если он имеется, кутикулы и кутикулярных слоев у клеток верхней кожицы обычно более значительна, чем у клеток нижней кожицы. Волоски у опушенных листьев располагаются преимущественно или даже исключительно на нижней стороне листа. Клетки кожицы, расположенные над проводящими пучками, особенно над армированными и над тяжами механических тканей, вытянуты в направлении, параллельном пучку или тяжу, и слабо извилисты. У многих злаков кожица состоит из клеток
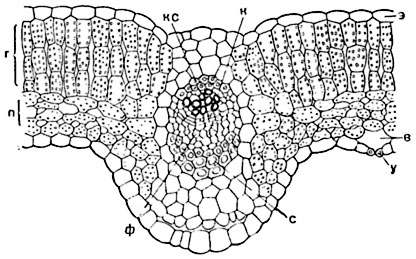
Рис. 174. Средняя часть поперечного разреза листа редьки (Raphanus sativus):
э - верхний эпидермис (верхняя кожица); у - устьице в нижней кожице; в - воздухоносная полость ("дыхательная полость"); г - палисадная ткань; п - губчатая ткань; в центре - сосудисто-волокнистый пучок; кс - ксилема; к - камбий; ф - флоэма; с - склеренхима.
199
нескольких типов, сильно отличающихся друг от друга. Из идиобластов в кожице листа отметим опорные клетки и цистолиты. У некоторых папоротников над жилками некоторые клетки эпидермиса, расположенные в ряд или одиночные, превращены в весьма толстостенные, теряющие живое содержимое паренхимные или прозенхимные стереиды. У некоторых крапивоцветных отдельные клетки содержат цистолиты.
У большинства наших деревьев и кустарников устьица образуются только на нижней стороне листа.
У наземных трав устьица имеются в большинстве случаев и на нижней и на верхней стороне листа, но на верхней в меньшем количестве (рис. 86).
Устьиц нет в верхней кожице у большинства растений с плотными кожистыми листьями и у растений с очень тонкими нежными листьями. У растений с плавающими на воде листовыми пластинками устьица находятся в верхней кожице. У растений, живущих в воде или приуроченных к глубокой тени, устьиц иногда мало или их не образуется совсем.
Частота устьиц - число их на 1 мм2 - у большинства растений колеблется от 40 до 300: у пшеницы, например, 50-70, у яблони и сливы - 250.
У листьев, вытянутых в длину (например, у злаков), устьица располагаются обычно продольными рядами, причем большая ось устьичной щели параллельна продольной оси листа. По поверхности листа устьица распределены неравномерно: наиболее тесно располагаются они в средней части листовой пластинки. Над тяжами механических тканей устьиц не образуется.
У некоторых растений под кожицей находятся слои бесцветной гиподермы - подкожной ткани, обычно водозапасающей, реже механической.
Мезофилл. Мезофилл (ассимилирующая ткань, мякоть листа) у растений с тонкими листовыми пластинками состоит из тонкостенной зеленой паренхимы (хлоренхимы), иногда включающей вместилища выделений. У большинства двудольных хлоренхима дифференцирована на палисадную (столбчатую) ткань и губчатую (рыхлую) ткань (рис. 174, 178). Обычно к верхней кожице примыкает палисадная ткань, а к нижней - губчатая. Наиболее типичны клетки наружного слоя палисадной ткани. Они расположены перпендикулярно к кожице, длинны, узки и богаты хлорофилловыми зернами. Находящиеся глубже слои палисадной ткани состоят из клеток более широких, коротких и менее богатых хлоропластами. Нередко группы из 2-3 клеток палисадного слоя примыкают поперечными перегородками к одной клетке глубже лежащего слоя, а наиболее глубоко расположенные палисадные клетки - к верхним клеткам губчатой хлоренхимы,- к собирательным клеткам. Своеобразная форма и расположение клеток палисадной ткани объясняются как черты приспособления к отведению продуктов фотосинтеза2 во флоэму по пути, возможно более короткому и представляющему наименьшее сопротивление, и к снабжению палисадной ткани водой из ксилемы проводящих пучков.
Губчатая ткань состоит из 2-7 слоев клеток округлого или чаще извилистого очертания. В ней обычно сильно развита система межклетников, пронизывающих губчатую ткань по всем направлениям. Размеры воздухоносных пространств и длина выступа клеток, которыми клетки примыкают друг к другу, наиболее значительны в плоскостях, параллельных поверхности листа. Рыхлая хлоренхима обычно превышает столбчатую по суммарной толщине, но уступает ей по количеству хлоропластов и содержанию хлорофилла. В листе клещевины, например в палисадных клетках, насчитывали в среднем по 36 хлорофилловых зерен, а в клетках губчатой ткани, больших по объему, чем клетки палисадной ткани, только по 20. Отношения
200
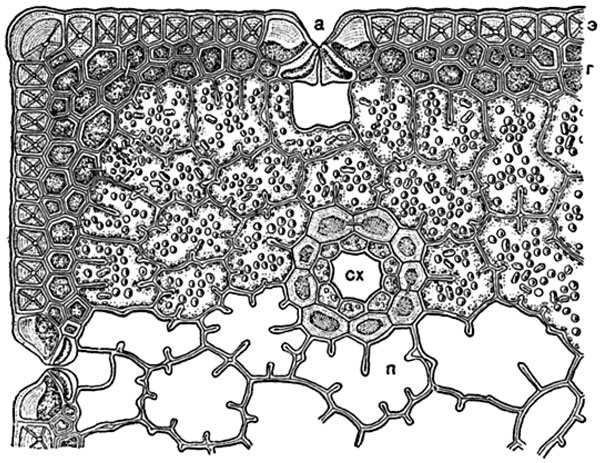
Рис. 175. Часть поперечного разреза хвои сосны (Pinus laricio):
э - эпидермис с сильно утолщенными клеточными стенками, в которых видны поровые каналы; г - гиподерма; а - углубление, под которым находится устьице; п - складчатая паренхима с хлорофилловыми зернами (не изображенными в некоторых клетках); сх - смоляной ход, окруженный эпителием и влагалищем из толстостенных клеток.
суммарного числа хлорофилловых зерен палисадной и губчатой паренхим на 1 мм2 поверхности листа изменяются приблизительно от 2 :1 до 6 : 1.
На сложение типичной губчатой паренхимы сильный отпечаток накладывает функция транспирации - отдачи воды в виде паров в атмосферу. Функция транспирации в значительной мере определяет строение губчатой паренхимы. Наличие межклетников надо считать соответствующей приспособительной чертой. Близ границы между палисадной и губчатой тканями располагаются мелкие проводящие пучки. В местах прохождения крупных пучков, особенно армированных, хлоренхима прерывается.
Особенности строения кожицы и мезофилла листа приводят к тому, что верхняя сторона листа обычно ярко-зеленая, нижняя же, вследствие меньшего содержания хлорофилла и обилия воздухоносных межклетников, более бледная.
Разделение на столбчатый и губчатый мезофилл встречается не только у двудольных растений, его можно наблюдать у некоторых папоротникообразных, голосеменных и однодольных.
У некоторых растений оболочки клеток мезофилла образуют складки, вдающиеся внутрь полости клетки, что сильно увеличивает внутреннюю поверхность клеток без увеличения их объема; возрастает площадь постенного слоя протоплазмы, в котором располагаются хлорофилловые зерна, т. е. увеличивается ассимилирующая поверхность.
Если складки расположены более или менее равномерно по всем стенкам клеток (рис. 175), то мезофилл носит название складчатого (сосны). У некоторых лютиковых (ветреница Anemone) и злаков (бамбук Bambusa) складки находятся только на стороне, обращенной к верхнему
201
эпидермису, и клетки их мезофилла часто называют дланевидными; могут быть и другие случаи расположения складок.
У некоторых листьев зеленая паренхима на обеих сторонах пластинки, на морфологически верхней и морфологически нижней, сходна3. Это наблюдается у растений, листья которых расположены в вертикальной плоскости (у нарциссов). У таких листьев обычно сходны на обеих сторонах также кожица и устьичный аппарат.
У большинства однодольных и многих голосеменных растений мезофилл однороден, т. е. все его клетки имеют приблизительно одинаковое строение. Это может быть простой или недифференцированный мезофилл - ткань из паренхимных клеток, почти изодиаметрических (многие злаки, рис. 176), вытянутых вдоль пластинки (гладиолус) или поперек пластинки листа (ирис). Бывают случаи, когда все клетки мезофилла складчатые (сосна) или палисадные.
Проводящая система. Проводящая система в листьях представлена "жилками", или "нервами", крупными и мелкими. Типы жилкования: 1) одна или несколько жилок, идущих вдоль пластинки, не ветвящихся и не соединенных анастомозами, или перемычками (у многих из хвойных); 2) жилки ветвятся, но ответвления их не соединяются перемычками, а окончания самых мелких жилок примыкают к краю листовой пластинки (у большинства папоротников, у гинкго Ginkgo biloba) 4; 3) часто несколько жилок идут почти параллельными прямыми или дугами, которые сближены у основания и у верхушки и соединяются тонкими поперечными или косо направленными анастомозами (у большинства однодольных); 4) у двудольных обычно лист обладает одной крупной жилкой 1-го порядка (у перистонервных листьев) либо несколькими, более или менее равносильными, крупными жилками (у пальчатонервных листьев).
При жилковании 2-го и 4-го типов от жилок 1-го порядка отходят ответвления 2-го порядка, дающие в свою очередь ответвления 3-го порядка и т. д.5. Ответвления соединяются анастомозами, идущими по различным направлениям; некоторые ответвления последнего порядка заканчиваются слепо среди мезофилла (рис. 177). При нервации этого типа, называемой сетчатой, мезофилл при рассматривании листа в плане оказывается разбитым на мелкие поля - многоугольные участки, окруженные жилками. Обычно внутрь поля проникает слепо заканчивающееся мельчайшее ответвление проводящей системы. Более крупные жилки находятся в ребрах, выступающих на нижней стороне листа, а у некоторых растений и на верхней.
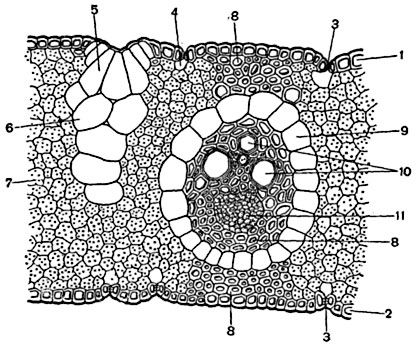
Рис. 176. Часть поперечного разреза листа злака Arundo donax:
1 - верхняя кожица; 2 - нижняя кожица; 3 - устьица; 4 - "дыхательная полость"; 5 - пузыревидные (водоносные) клетки кожицы; 6 - пузыревидные клетки основной ткани листа; 7 - клетки хлоренхимы (ассимиляционной ткани); 8 - склеренхима; 9 - эндодерма проводящего пучка; 10 - сосуды в ксилеме пучка; 11 - флоэма.
202
Мелкие жилки можно видеть, рассматривая пластинку листа в проходящем свете; самые мелкие обнаруживаются только под микроскопом. Наиболее крупные из жилок содержат несколько проводящих пучков, нередко снабженных обложками из склеренхимы или из внутренней колленхимы, и, кроме того, паренхимную ткань. Армированными могут быть и некоторые из ответвлений жилок.
Проводящие пучки в листьях, как правило, коллатеральные. Ксилема находится в морфологически верхней, флоэма - в нижней части пучка. Пучки в пластинках листьев большей частью закрытые. У немногих трав и у древесных растений более крупные проводящие пучки открытые, но камбий в них функционирует слабо. У видов, обладающих биколлатеральными пучками в стеблях, биколлатеральны и более крупные проводящие пучки листьев. Сосудистые пучки не соединяются с мякотью листа непосредственно, а всегда бывают окружены, как чехлом или футляром, клетками обкладки - одним слоем тонкостенных, плотно сомкнутых паренхимных клеток (рис. 176, 177). Эти клетки вытянуты вдоль жилок, не содержат хлорофилловых зерен. Опыты показали, что продукты ассимиляции из губчатого мезофилла поступают в клетки обкладки и затем уносятся по ним до флоэмы более крупных жилок.
При рассматривании проводящих пучков (от более крупных к мелким) можно видеть, что пучки постепенно уменьшаются в поперечном сечении и упрощаются. Сначала отпадает арматура, затем уменьшается флоэма и меняется ее характер: ситовидные трубки заметно сужаются, тогда как клетки-спутницы или уменьшаются, но в меньшей мере, или даже приобретают большие поперечные размеры, чем в крупных пучках. Более тонкие пучки содержат ситовидные трубки с неясно выраженными ситечками или вовсе лишены их. Далее флоэма представлена лишь сравнительно крупными клетками-спутницами, или, точнее сказать, материнскими клетками, не поделившимися на ситовидные трубки и сопровождающие клетки. В мелких ответвлениях проводящей системы листа флоэма вовсе отсутствует. В самых мелких сосудах сильно упрощается и ксилема: трахей нет, число трахеид постепенно убывает, и последние ответвления проводящей системы заканчиваются единичными трахеидами, окруженными обычно обложкой из одного слоя клеток без межклетников и вытянутыми в направлении, параллельном жилке (рис. 177).
Механическая ткань. Механическая ткань (арматура) в листе двудольных представлена склеренхимой и колленхимой, однодольных и голосеменных - склеренхимой.
Склеренхима располагается в листе обычно при проводящих пучках только под флоэмой или в виде двух тяжей под флоэмой и над ксилемой (рис. 174), либо в виде сплошной обложки (рис. 176). Нередко склеренхима в мезофилле представлена еще тяжами или пластинками, примыкающими к кожице (рис. 178). Склеренхимные тяжи под кожицей находятся обычно напротив проводящих пучков и нередко смыкаются с их склеренхимными
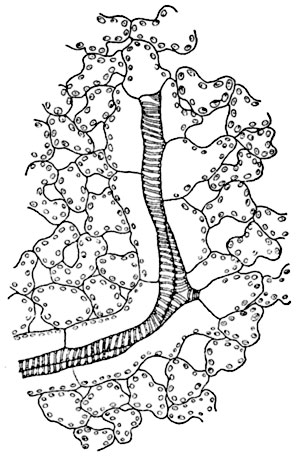
Рис. 177. Окончание проводящего пучка в листе недотроги мелкоцветной (Impatiens parviflora).
203
обложками. Гиподермальная склеренхима может представлять, как у некоторых хвойных, сплошную обложку, прерывающуюся лишь напротив устьиц и смоляных ходов (рис. 175). У двудольных колленхима обычно располагается под кожицей в виде тяжей - напротив проводящих пучков. У многих двудольных, в числе их у подорожников (Plantago), у борщевиков (Heracleum), обложки при пучках состоят из внутренней колленхимы. В крупных листьях вдоль края располагаются арматурные тяжи, находящиеся непосредственно под кожицей или несколько глубже. На дне вырезок у лопастных листьев нередко находятся крупные изогнутые стереиды, предохраняющие листовую пластинку от надрывов. У многих растений (например, у смородин) в листьях имеются анастомозы пучков, проходящие в виде дуг, касательных к дну выемок края листа. Противодействует механическим повреждениям пластинки и кожица, являющаяся прочным и упругим футляром листа; по краям пластинки и в глубине выемок листа клетки кожицы имеют особенно сильно утолщенные наружные стенки. У многих растений прочность листа и сохранение им определенного положения в пространстве обеспечивается тем, что листовая пластинка в целом (у некоторых злаков) либо доли или лопасти ее (у многих пальм) имеют на поперечных разрезах глубоко-желобовидную форму (в виде букв V или W).
У некоторых растений (чай, маслина), кроме механических тканей, в листе встречаются и отдельные механические клетки - склереиды, иногда очень крупные (рис. 79).
Лист представляет собой структуру, очень сильно изменяющуюся от влияния внешних условий: количества почвенной и дождевой влаги, особенностей почвы, света, ветра, температуры, положения на стволе и т. п. Несмотря на то что общий план строения листа одинаков, в деталях оказывается много вариантов. Наиболее характерными будут изменения, вызванные влиянием света и положением листа на стебле (его ярусностью).
Световые и теневые листья древесных растений. У деревьев и кустарников листья, находящиеся при полном зазеленении кроны в условиях более яркого освещения (световые листья), отличаются по строению от затененных, находящихся внутри кроны или в нижней ее части (теневых) листьев. Листья световые (расположенные на периферии кроны) даже внешне отличаются от теневых большей толщиной и жесткостью. Во внутреннем строении световых и теневых листьев также отмечаются большие различия. По внутреннему строению световые листья отличаются от теневых следующими особенностями. Клетки кожицы их имеют менее извилистые очертания и более толстостенны; они относительно беднее хлорофиллом; число устьиц на единицу поверхности листа более высоко; ассимиляционная ткань, особенно палисадная, более мощна. Например, у бука световые листья имеют два слоя палисадных клеток, высоких, узких и тесно расположенных, и 3-4 слоя губчатой паренхимы, теневые же - один слой палисадной и три слоя губчатой ткани (рис. 178, 179).
Характер структуры листа, теневой или световой, зависит не только от освещенности листа после выхода его из почки, но и от интенсивности освещения почки, в которой лист заложился и прошел первые фазы формирования. Это было выяснено опытами такого рода. Различные побеги бука, срезанные до распускания почек, ставились до полного распускания листьев в воду и выдерживались в тени и на солнце. В обоих вариантах освещения листья почек побегов, взятых из световой части кроны, развивались в листья со световой структурой, листья же почек теневых побегов - в листья теневой структуры6.
204
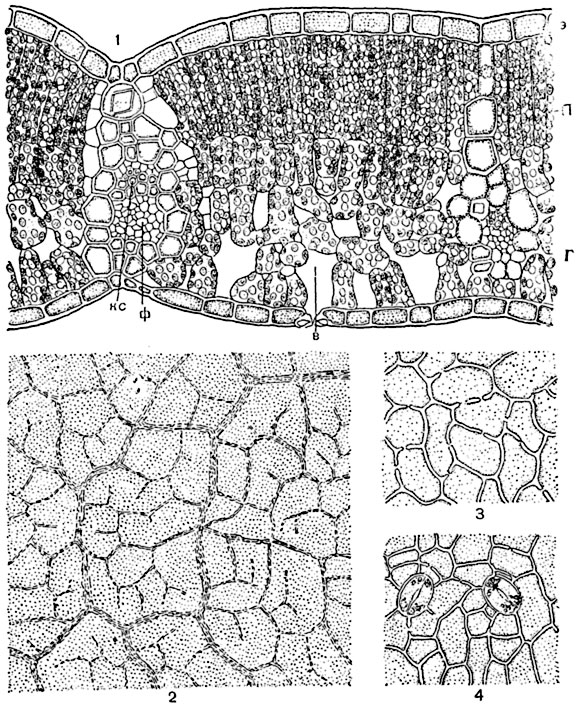
Рис. 178. Световой лист бука (Fagus silvatica):
1 - поперечный разрез части листовой пластинки: э - кожица, п - палисадная ткань, г - губчатая ткань, в - воздухоносная полость над устьицем ("дыхательная полость"), кс - ксилема проводящего пучка, ф - флоэма его; 2 - вид участка листа при малом увеличении, в плане, просвечивает сеть проводящих пучков с их окончаниями; 3 - участок верхней кожицы, в плане; 4 - участок нижней кожицы с устьицами, в плане; фиг. 1, 3, 4 - при большом увеличении, фиг. 2 - при малом.
И. Г. Серебряковым были поставлены (1944-1945) опыты в более естественных условиях над взрослым деревом черемухи. В конце апреля 1944 г., до раскрытия почек, над световыми ветками был натянут зонт из плотной зеленой ткани. Опыт продолжался до лета 1945 г. В строении листьев на световых ветках, ставших затененными, наметился сдвиг в сторону теневой структуры, который стал более значительным в следующем году.
Не все черты строения листа одинаково изменяются при экспериментальном воздействии. Выяснилось, что мощность развития палисадной ткани определяется условиями освещения, а устьичного аппарата и жилкования - условиями водного режима.
Формирование листьев по световому или теневому типу зависит также и от состояния организма, в частности от возраста всего растения и от возраста данного побега: молодые деревца и кустарники несут листья лишь теневого типа; способность образовывать листья теневые и световые
205
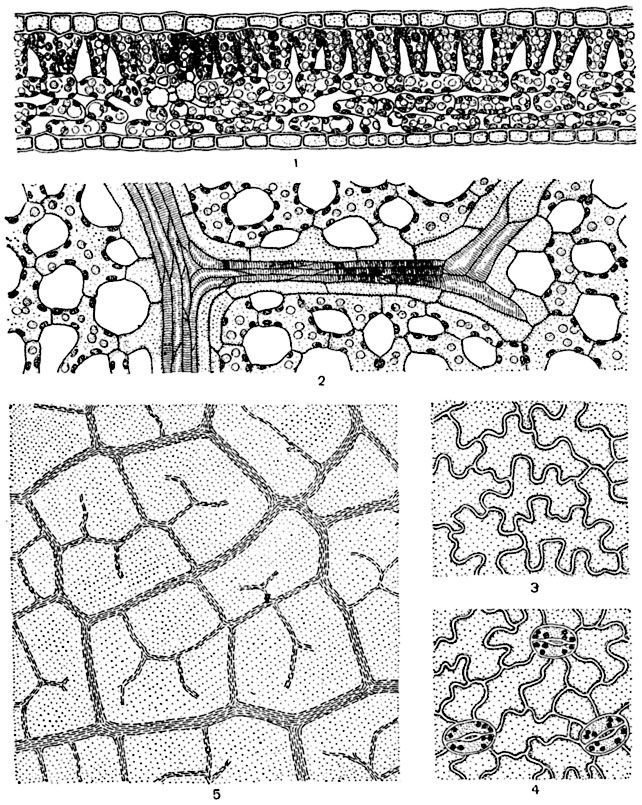
Рис. 179. Теневой лист бука:
1 - частичный поперечный разрез листовой пластинки: палисадная ткань из одного слоя, губчатая - из трех слоев клеток; 2 - часть продольного среза, проходящего параллельно поверхности листовой пластинки, через губчатую ткань; 3, 4 и 5 аналогичны 2, 3 и 4 рисунка 178; 1, 2, 3 и 4 - при большом увеличении, 5 - при малом.
проявляется в более зрелом возрасте. На побегах, развивающихся из спящих глазков или из придаточных почек, сначала образуются листья теневого типа и лишь по истечении нескольких лет могут появляться листья светового типа.
Образование листьев преимущественно или исключительно по теневому типу может быть вызвано не только продолжительным слабым освещением и увлажнением воздуха, но и сильной обрезкой, удобрением почвы.
Строение листьев растений, живущих в особых условиях водоснабжения, температуры, освещения и т. д., имеет свои особенности, подробно излагаемые в курсах физиологии и экологии растений.
Закон В. Р. Заленского. Структура взрослых листьев данного побега имеет некоторые различия в зависимости от ярусности их - положения на
206
стебле. Закономерности в этих различиях были четко и подробно сформулированы впервые Вячеславом Рафаиловичем Заленским (в 1902 и 1904 гг.) и известны в науке под именем "закона Заленского". Этот закон выражен его автором в применении к травянистым растениям в виде следующих положений: суммарная длина жилок листа на единицу поверхности листовой пластинки возрастает от нижнего яруса листьев к верхнему, что наиболее резко выражено у растений сухих и солнечных местообитаний. Переходя от листа к листу в направлении от нижнего яруса - близ основания стебля - к верхнему, можно заметить, кроме того, что более высоко расположенные листья имеют меньшие размеры клеток верхней и нижней кожицы, волосков (если они имеются), замыкающих клеток устьиц, клеток мезофилла, а также более слабо развитую систему, межклетников. Извилистость боковых стенок клеток кожицы листьев понижается у выше сидящих листьев по сравнению с ниже сидящими. От яруса к ярусу - в направлении снизу вверх - увеличивается в листьях частота устьиц и волосков, мощность воскового налета, толщина наружных стенок клеток кожицы и относительная мощность арматуры. Палисадная ткань развивается тем типичнее, а губчатая ткань тем слабее, чем выше на стебле расположен лист.
Различия в строении листьев различных ярусов отчасти объясняются тем, что листья, расположенные выше, находятся в условиях худшего снабжения водой и большей транспирации. Таким образом, листья верхних ярусов оказываются как бы более "ксероморфными".
Закон Заленского в известной мере применим к растениям одного и того же вида, но различных местообитаний (например, для растений, растущих вблизи реки и на водоразделе).
Бывают случаи, когда ярусные различия не проявляются; это также связано с условиями произрастания и особенностями растения.
Например, у лиан сырого и сильно затененного местообитания различия в строении между листьями разных ярусов очень слабо выражены; у плющей - обыкновенного (Hedera helix) и колхидского (H. colchica) - даже обнаружены отношения обратного, нежели следовало бы по закону Заленского, характера. Это объясняется наличием у плющей придаточных воздушных корней: листья верхних ярусов могут снабжаться водой не хуже или даже лучше, чем листья нижних ярусов.
207
1 От греческого "мезо" - средний и латинского phyllum - лист.
2 Быстрое отведение из листа продуктов ассимиляции благоприятствует ходу фотосинтеза.
3 Листья с таким строением называют нередко изолатеральными; этот термин неудачен, лучше называть их эквифациальными.
4 Ветвление жилок у Ginkgo нередко дихотомическое (вильчатое).
5 У некоторых растений можно найти жилки 7-го и даже 8-го порядков.
6 Под световыми и теневыми побегами разумеются побеги, ярко либо слабо освещаемые после полного зазеленения кроны. После зазеленения кроны начинается заложение листьев будущего года и проходят первые этапы их развития.