

Перекрестное опыление
Анемофилия. Анемофильными являются, по-видимому, около 1/10 всех покрытосеменных растений. К ним относятся почти все злаки, осоковые, большинство пальм, большинство наших деревьев (березы, ольхи, осины, тополя, вязы, дубы, бук, грецкий орешник, орешник-лещина, шелковицы, платаны и др.), хмель, конопля, крапивы, подорожники, рдесты и др. Цветки у них обычно мелкие, невзрачные, с простым чашечковидным околоцветником, без ярко окрашенного венчика. Пыльца сухая, мелкая, развивающаяся в огромных количествах. Например, в одной сережке орешника-лещины около 4 млн. пылинок, в мужской метелке кукурузы - около 50 млн. пылинок. У некоторых анемофильных растений, например у крапив, тычиночные нити, закрученные в бутоны, при раскрывании цветка с силой раскручиваются и разбрасывают пыльцу из лопнувших пыльников (рис. 305, 4). У злаков, конопли, щавеля, василистников, подорожников
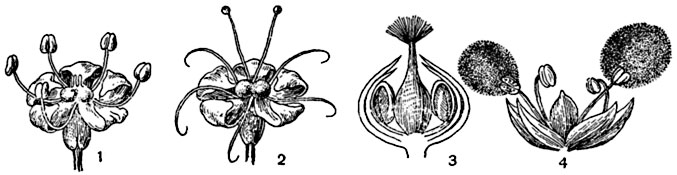
Рис. 305. Дихогамия (неодновременное созревание тычинок и пестиков):
1-2 - протерандричный цветок бедренца (Pimpinella saxifraga); 3-4 - протерогиничный цветок стенницы (Parietaria officinalis).
и др. пыльники сидят на длинных, высовывающихся из цветка, легко раскачиваемых тычиночных нитях. У многих мужские соцветия повислые, легко раскачиваемые сережки (березы, орешники, осины, тополя, дубы и т. п.). Улавливание пыльцы у анемофильных растений производится длинными, высовывающимися из цветков, волосистыми, перистыми рыльцами (злаки, многие древесные породы и др.). Растут анемофильные растения большей частью большими зарослями из одного или немногих видов. Цветение большинства лиственных анемофильных древесных пород ранней весной, до или одновременно с распусканием листьев, облегчает улавливание пыльцы рыльцами. Самоопылению у анемофильных растений препятствует одно- или двудомность их или наблюдающееся у многих неодновременное созревание пыльников и рылец.
О том, на какие расстояния может переноситься пыльца, могут дать представление следующие наблюдения. У берегов Скандинавии ловили пыльцу хвойных, березы и др. на судах, находившихся за 30-55 км от берега. На Фарерских островах, на Новой Земле находили пыльцу орешника, ольхи, березы и других растений, которая могла быть занесена туда
360
только из мест, отстоящих на 400 км. Эти цифры еще не говорят о том, что эта пыльца была способна к прорастанию и оплодотворению. Но, например, в Италии, близ Отранто, женский экземпляр финиковой пальмы был оплодотворен пыльцой, которая могла быть занесена туда с мужского экземпляра, росшего за 64 км.
Энтомофилия. Для энтомофильных растений характерен ярко окрашенный венчик или венчиковидный околоцветник, делающий их издали заметными для насекомых. В случае мелких цветков они обычно бывают собраны в крупные, хорошо заметные соцветия (у зонтичных, мареновых, сложноцветных и др.). Цветки, находящиеся на периферии соцветий у многих сложноцветных (маргаритки, ромашки, васильки и т. п.), у калины и др., бывают крупнее центральных; у некоторых (васильки, калина и др.) они при этом имеют лишь редуцированные тычинки и пестики или совсем не имеют их, являются бесплодными, и, таким образом, в соцветии происходит разделение функций: краевые бесплодные цветки привлекают насекомых, опыляющих срединные, более мелкие цветки и невзрачные, но приносящие плоды. В соцветиях многих зонтичных (морковь, борщевик и др.) и скабиоз сильнее развиты даже не целиком все краевые цветки, а лишь те лепестки их, которые находятся на наружных сторонах соцветия и обрамляют его со всех сторон. У некоторых сложноцветных (ромашки, маргаритка и др.) краевые цветки корзинки иначе окрашены, чем центральные, и благодаря такой контрастности соцветие становится еще заметнее.
У некоторых (например, у посевной вики) лепестки вначале бывают неодинакового цвета; гораздо чаще контрастность окраски получается вследствие различных, иначе окрашенных полос, пятен и т. п., делающих венчик или венчиковидный околоцветник узорчато разрисованным.
У некоторых растений различных семейств (синеголовника, шалфеев Salvia splendens, sclarea, молочаев и др.) бывают окрашены в другой цвет, чем венчик, не только цветки, но и верхушечные листья в соцветиях (кроющие и др., например, у иван-да-марьи) и даже оси соцветий (бегонии и др.). Корзинки некоторых сложноцветных хорошо заметны благодаря яркой окраске листочков их обертки (так называемые бессмертники, кошачья лапка и др.). Об окраске чашечки уже упоминалось на странице 329, о многочисленных окрашенных тычинках, заменяющих функционально венчик, - на странице 335.
Кроме яркой окраски цветков, большое значение имеют и различные запахи их, зависящие от выделения эфирных масел, издалека действующих на обоняние насекомых и ориентирующие их в разыскивании цветков1. Число различных запахов, выделяемых цветками, исчисляется сотнями. Ряд растений имеет цветки с отвратительными запахами, напоминающими запахи трупов, тухлой рыбы, разлагающейся мочи, навоза и т. п. (многие раффлезиевые, стапелии, некоторые тропические орхидеи и др.). Цветки эти нередко напоминают по окраске разлагающееся мясо и посещаются мухами и жуками, водящимися на падали и испражнениях.
Окраска и запах лишь указывают насекомому, куда ему лететь. Посещают же они цветки ради нектара и отчасти пыльцы, служащих им пищей. У сравнительно очень немногих растений насекомые, производящие перекрестное опыление, поедают также сочные волоски на тычиночных нитях (коровяк, традесканция и др.) и других частях цветка, бородавочки и т. п. Пыльца служит единственной приманкой для насекомых у меньшинства энтомофильных растений; в среднеевропейской флоре к ним
361
относят около 10% из всех опыляемых насекомыми. Цветки у многих из них крупные, открытые, актиноморфные, не поникающие; тычинки многочисленные, далеко выступающие, дающие массу пыльцы. Таковы маки, шиповники, ветреницы (Anemone, Hepatica), мимозы, отчасти коровяки и др. Пыльца поедается насекомыми, а также, как известно, в больших количествах собирается пчелами, шмелями и откладывается в сотах как запас пищи для личинок (так называемая перга, или хлебина пчеловодов).
В строении околоцветника, расположении пыльников, рылец и т. п. наблюдается невероятное количество разнообразнейших приспособлений к форме тела и повадкам насекомых, производящих у них перекрестное опыление. На венчике или венчиковидном околоцветнике бывают часто различные выросты, бугорки, волоски и т. п., расположенные так, чтобы насекомое, обходя их, направлялось к нектару по тому пути, где оно должно коснуться тычинок и рыльца. Этой же цели, быть может, служат различные узоры, иначе окрашенные штрихи, пятна на венчике или венчиковидном околоцветнике, сходящиеся обычно к тому месту, где находятся нектарники. Их называют указателями нектара, хотя весьма возможно, что во многих случаях эта роль их очень преувеличивается.
У большинства энтомофильных растений главной приманкой для насекомых является нектар, хотя, конечно, у многих из них поедается частично и пыльца. Нектарники обычно расположены так, что, добираясь до них, насекомое касается тычинок и рыльца.
Пыльца энтомофильных растений обычно отличается от пыльцы анемофильных неровной поверхностью, покрытой шипиками, бородавочками и другими выростами, способствующими переносу ее насекомыми; у некоторых она бывает клейкая. Размеры пылинок у энтомофильных растений обычно несколько крупнее, чем у анемофильных.
Растения, у которых перекрестное опыление производится только одним, определенным насекомым, очень немногочисленны. Обычно в зависимости главным образом от расположения нектарников цветки определенного строения могут опыляться теми или иными группами насекомых.
Насекомые с коротким хоботком, как мухи и другие двукрылые, жучки и т. п., могут опылять только цветки с лежащими открыто, легко доступными нектарниками, Это наименее специализированные из энтомофильных цветков, преимущественно актиноморфные, с преобладающими белыми, желтыми, зеленовато-желтыми, реже розовыми и красноватыми окрасками. Многие имеют неприятный запах и опыляются мухами, в том числе и теми, которые откладывают яички в навоз, падаль. Сюда относятся многие зонтичные, камнеломковые, крушина, бузина, подмаренники, клены, многие розоцветные и т. д.
Цветки с более глубоко скрытым нектаром могут опыляться насекомыми с более длинным хоботком - пчелами, шмелями, осами и другими перепончатокрылыми. У многих из них цветки зигоморфные, окраски преобладают голубые, синие, фиолетовые, пурпуровые2. К этой группе относятся многие мотыльковые, губоцветные, норичниковые и др.
Наконец, цветки с нектаром, скрытым на дне длинных, узких трубочек венчика или в шпорах, могут опыляться только бабочками, имеющими очень длинный хоботок, в 10-80 и более (до 300) миллиметров. Цветки, опыляемые дневными бабочками, бывают часто оранжевого или чисто красного цвета, ночными - белого, светло-розового, светло-желтого с более или менее сильным запахом, который у многих становится сильнее к вечеру. К этой группе относятся табаки, многие гвоздичные, многие орхидные (ятрышники, белая ночная фиалка и др.), крокусы, нарциссы, каприфоль и др. У некоторых, опыляемых сумеречными и ночными бабочками, цветки раскрываются только на ночь, а на день закрываются или к утру уже увядают (каперцы, белый душистый табак и др.).
Само собой разумеется, что между указанными группами имеются многочисленные переходы и что насекомые с длинным хоботком часто посещают и опыляют и цветки с открыто лежащим нектаром.
Кроме того, следует заметить, что некоторые виды (например, многие вересковые) в различных местностях и даже в одной и той же местности в разных экологических условиях в разные годы могут опыляться различно - то энтомофильно, то анемофильно.
362
Вопросы перекрестного опыления могут иметь и чисто практическое значение. Урожай плодов и семян энтомофильных культивируемых растений падает, если в период цветения их стоит дождливая или холодная погода, неблагоприятная для лёта насекомых-опылителей. Наоборот, урожай повышается на 40-50%, а в некоторых случаях даже в 3-4 раза, если вблизи от данной культуры находится пасека с пчелами или если ульи вывозятся на соответствующее поле.
В последнее время нередко практикуется направленное пчелоопыление, или "дрессировка" пчел; она имеет целью приучить пчел посещать те цветки, которые они мало посещают, но опыление которых пчелами важно для человека (например, для получения семян красного клевера, опыляемого в природе шмелями и мало посещаемого пчелами). При такой "дрессировке" в улей ежедневно ставят для подкормки пчел небольшие порции сахарного сиропа, в который кладут цветки, к посещению которых хотят приучить пчел. Привыкая к их аромату, пчелы увеличивают посещения желаемых растений нередко в 10-15 раз.
Орнитофилия. В тропиках цветки многих растений опыляются мелкими птичками (колибри, медососы и др.), питающимися их нектаром. Для многих орнитофильных цветков характерна ярко-красная окраска, которая, по-видимому, лучше всего различается птицами. Птицы могут некоторое время, не садясь на цветки, держаться в воздухе и высасывать нектар. Пыльца откладывается обычно на головные перья.
Гидрофилия. К гидрофильным растениям относятся такие, у которых все растение, в том числе и цветки, целиком погружено в воду. Сюда принадлежат, например, роголистники (Ceratophyllum), морская трава, или взморник (Zostera), наяда (Najas) и некоторые другие. Пыльца у них не имеет экзины, защищающей обычно ее от высыхания. Пылинки, переносимые пассивно водой, имеют часто нитевидную форму, лучше обеспечивающую возможность соприкосновения их с рыльцами. Последние также нередко имеют удлиненную нитевидную или лентовидную форму, тоже лучше обеспечивающую улавливание пыльцы. У многих удельный вес пыльцы одинаков с удельным весом воды, так что пыльца не всплывает и не тонет и переносится водными течениями. У некоторых растений, имеющих женские цветки, расположенные ниже мужских, пыльца тяжелее воды и при медленном опускании ее вниз попадает на рыльца. Между надводным и подводным опылением имеются переходы. Подводное опыление следует считать выработавшимся в ходе эволюции растений из надводного.
Дихогамия. Перекрестному опылению содействует чрезвычайно широко распространенное явление неодновременного созревания тычинок и рыльца, носящее название дихогамии3. Раннее вскрывание пыльников, когда рыльцев цветке еще не готово к восприятию пыльцы, называется протерандрией4 или протандрией (рис. 305, 1, 2); более раннее созревание рыльца при еще не вскрывшихся пыльниках - протерогинией5 или протогинией (рис. 305, 3, 4).
Дихогамия наблюдается не только в обоеполых, но и в однополых цветках одно- и двудомных растений, у которых раньше вскрываются или тычиночные, или пестичные цветки.
Протерандрия встречается у гораздо большего числа растений, чем протерогиния. Быть может, это нужно поставить в связь с тем, что тычинки, расположенные в цветке ниже пестиков, закладываются и начинают развиваться раньше их. Протерандрично большинство гвоздичных, гераниевых, мотыльковых, мальвовых, губоцветных, колокольчиковых, сложноцветных, многие лилейные, свекла, кукуруза, наперстянка и др. Протерогиния наблюдается у большинства крестоцветных, розоцветных, пасленовых,
363
жимолостных, зимовника, душистого колоска, ситников, большинства одно- и двудомных растений и т. д. У многих рыльца высовываются из еще не раскрывшегося околоцветника.
Среди семейств и родов протерандричных бывают роды и виды протерогиничные, и наоборот. Имеются также наблюдения, что одно и то же растение может быть в одних местностях протерандричкым, в других - протерогиничным, в третьих - гомогамным6, т. е. с одновременно созревающими пыльниками и рыльцами.
Гетеростилия. Существуют растения, у которых одни экземпляры имеют цветки с длинными столбиками, другие - с короткими. Соответственно этому расположены и пыльники: у одних - ниже, у других - выше собственного рыльца, таким образом, что они всегда приходятся на уровне, соответствующем положению рыльца в цветках другой формы (рис.306). Явление это носит название гетеростилии7 или разностолбчатости и наблюдается у первоцветов, проломников, турчи и многих других первоцветных, у трифоли (Menyanthes trifoliata), некоторых генциан, незабудок, медуниц (Pulmonaria) и еще некоторых бурачниковых, у гречих и т. д. (рис. 306). Гетеростилию можно рассматривать как защиту от так называемой гейтоногамии8, т. е. от опыления цветков данного экземпляра растения пыльцой с других цветков того же самого экземпляра. Перекрестное опыление пыльцой с других экземпляров, имеющих хотя бы минимальные отличия и выросших в несколько иных условиях, дает лучшие результаты и потомство, более разнообразное в своих приспособительных возможностях.
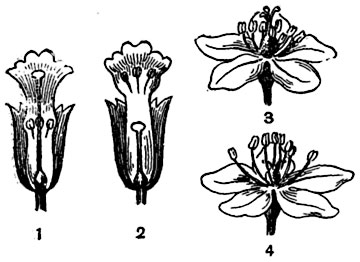
Рис. 306. Гетеростилия в цветках:
1-2 - первоцвет; 3-4 - гречиха.
Результаты опыления получаются лучшие, если пыльца из длинностолбчатых цветков попадает на рыльце короткостолбчатых, и наоборот (так называемое легитимное9 опыление). В противных случаях (так называемое иллегитимное10 опыление) семян или совсем не образуется (у медуниц), или их бывает меньше (у гречихи) и из них вырастают более слабые растения.
С гетеростилией связаны и известные приспособления пыльцы к рыльцу, и обратно. У форм с высоко расположенными пыльниками пыльца крупнее, чем в низко прикрепленных пыльниках11. У форм с длинным пестиком сосочки рыльца крупнее и дальше отстоят друг от друга, чем у короткостолбчатых: первые приспособлены к задержанию более крупной пыльцы.
364
Некоторые примеры более сложных приспособлений к перекрестному опылению
У шалфеев, имеющих всего две тычинки, связник их очень длинный и подвижно сочленен с тычиночной нитью, качаясь на ней вроде рычага или коромысла (рис. 307). Пыльца развивается только в двух гнездах пыльника на верхнем длинном конце связника; на нижнем же, коротком, из другой половины пыльника образуется небольшое лопатообразное расширение совсем без пыльцы или с малым количеством ее. Раньше обычно описывалось на основании старых наблюдений немецкого ботаника X. К. Шпренгеля, что опыление у шалфеев происходит путем прикосновения спинки шмеля к рыльцу, находящемуся на конце столбика, свисающего вниз в более старых цветках. Советский ботаник Н. Г. Холодный установил, что столбик свисает вниз в цветках шалфея лишь при увядании их после опыления, а опыление происходит иначе. Когда насекомое (шмель) просовывает хоботок внутрь цветка, то оно толкает короткие плечи коромысла связников; длинные плечи их при этом опускаются вниз, ударяют насекомое по спинке и высыпают на него пыльцу. Когда затем насекомое, посещая другие цветки шалфея, улетает с них, то от взмахов его крыльев подсохшая пыльца (из ранее посещенных цветков) поднимается со спинки в воздух и опыляет рыльце пестика, высовывающееся из-под верхней губы венчика.
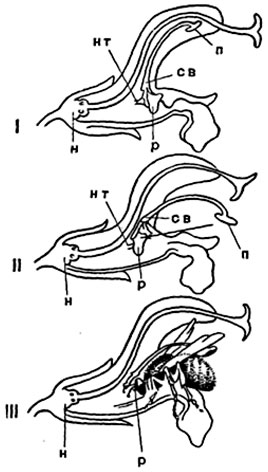
Рис. 307. Перекрестное опыление у шалфея:
н - нектарник; р - расширение; св - связник; нт - нить тычинки; п - ее пыльник; I и II - цветки до посещения; III - в момент посещения шмеля.
Большинство орхидных имеет всего одну тычинку, которая срастается нитью со столбиком пестика. Половинки пыльника, находящиеся над рыльцем, разъединены связником; в каждой половинке вся пыльца сливается в общую массу, так называемый поллиний12, от которого отходит ножка из затвердевшей слизи (рис. 308, 3). Под пыльником находится трехлопастное рыльце; две боковые лопасти его могут воспринимать пыльцу, а средняя превращена в мешочек, в котором лежат липкие подушечки; к последним прикреплены ножки, отходящие от поллиниев. Поллиний вместе с ножкой и подушечкой называют поллинарием. Когда насекомое всовывает головку в цветок, то к ней приклеиваются липкими подушечками поллинарии, вытаскиваются из цветка и переносятся на другое растение; при этом во время полета насекомого ножки поллиниев подсыхают, поллинии загибаются вниз и при посещении другого цветка приходятся как раз против воспринимающих их лопастей рыльца. Обстоятельные исследования о приспособлениях различных орхидных к перекрестному опылению были произведены Ч. Дарвином.
У кирказонов (Aristolochia clematitis и другие виды) простой околоцветник имеет вид трубки, расширенной у входа, суженной посредине и шаровидно раздутой у основания (рис. 309). В этом шаровидном вздутии помещаются тычинки и пестик. Пыльники срастаются спинками со столбиком и расположены внизу под шестилопастным рыльцем. Цветки протерогиничны. Мелкие комарики и мушки заползают в околоцветник и живут несколько дней внутри его нижнего шаровидного вздутия, так как им мешают выбраться наружу волоски в узкой части околоцветника, направленные
365
внутрь. Они питаются здесь сочными клетками на внутренних стенках околоцветника. Когда вскроются пыльники, волоски в узком проходе околоцветника вянут; насекомые, обсыпанные пыльцой, могут выбраться из цветка и, перелетев на другие, позднее раскрывшиеся цветки, опылить их рыльце.
Аналогично кирказонам происходит перекрестное опыление у аронников (Arum). У последних все соцветие играет ту же роль, что околоцветник кирказонов. Оно имеет вид початка, окруженного крупным, у многих лепестковидно окрашенным верхушечным листом, так называемым чехлом или крылом (рис. 310). Самая верхняя часть початка не несет цветка; пониже ее расположены мужские цветки, а еще ниже, после небольшого промежутка, женские. Растение протерогинично. Мелкие мушки заползают внутрь чехла и в течение некоторого времени не могут выбраться наружу, так как им мешают многочисленные волоски, расположенные на оси початка и направленные вниз. Вследствие энергичного дыхания цветков температура внутри чехла поднимается выше температуры окружающего воздуха. Когда вскроются пыльники, волоски вянут; мушки, обсыпанные пыльцой, могут выбраться наружу и, попав в другое соцветие, опылить его рыльца.
Очень оригинально происходит опыление у двудомной валлиснерии, разводимой часто в аквариумах и дикорастущей у нас, укореняясь на дне водоемов, на юге Европейской части Союза, кое-где в Средней Азии, на Дальнем Востоке. Женские цветки ее сидят на цветоножках, первоначально спирально закрученных; затем цветоножки раскручиваются и выносят цветки на поверхность воды, где те раскрывают околоцветник и три бахромчатых, высовывающихся из него рыльца. Мужские цветки около 0,5-1 мм в диаметре, образующиеся тоже под водой, отрываются от цветоножек, всплывают на поверхность, где раскрываются и плавают (рис. 311). Подгоняемые ветром, они подплывают к женским цветкам и опыляют их липкой, не сдуваемой ветром пыльцой. После этого цветоножки женских цветков снова скручиваются спирально и втягивают цветки под воду, где происходит развитие плодов и семян.
У инжира, или винной ягоды (Ficus carica), мелкие цветки находятся на внутренней поверхности мясистого, грушевидного, полого внутри вместилища, образованного сросшимися веточками так своеобразно измененного соцветия. На одних экземплярах в этих вместилищах находятся только женские цветки, имеющие
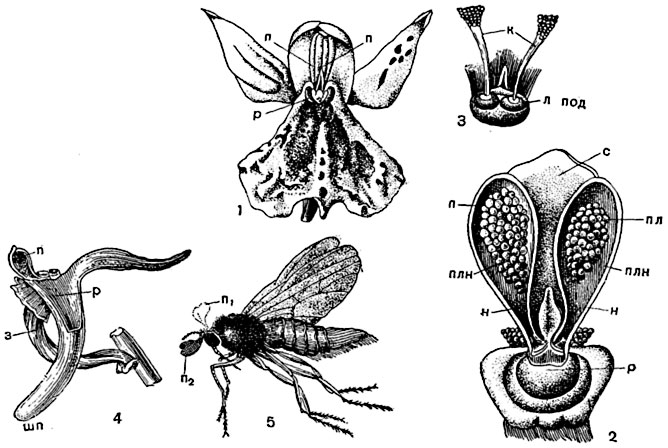
Рис. 308. Опыление у орхидных:
1 - цветок ятрышника (вид спереди): 2 - рыльце и тычинка; 3 - ножки поллиниев с липкими подушечками; 4 - вид цветка сбоку с удаленным околоцветником; 5 - насекомое с поллинариями орхидного: п1, п2 - первоначальное положение поллинариев; пл - поллиний; н - ножка поллиния; л под - липкая подушечка; п - поллинарий; з - завязь; р - рыльце; плн - половинка пыльника; с - связник; шп - шпорец.
366
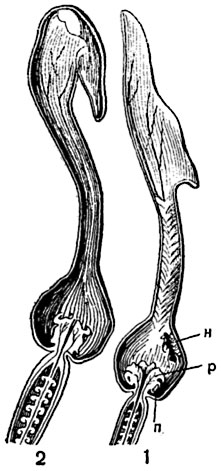
Рис. 309. Продольный разрез молодого (1) и старого (2) цветков кирказона:
п - пыльники; р - рыльце; н - насекомое.
пестик с длинным столбиком. На других экземплярах, в верхней части вместилища, находятся мужские цветки, а ниже их - женские с короткими столбиками (рис. 312). Опыление производится очень маленькой осой бластофагой (2 мм длины). Она откладывает яйца (по одному) в короткостолбчатые женские цветки, завязь которых после этого сильно разрастается и служит приютом и пищей личинке бластофаги. Когда из личинок разовьются взрослые насекомые, у них здесь же происходит оплодотворение, после чего самцы умирают, не вылетая из соцветия, а самки покидают вместилища и при этом перепачкиваются пыльцой расположенных у его отверстия мужских цветков. Они заползают в другие, позже развившиеся соцветия (цветение происходит несколько раз в году) и откладывают там яички. Если осы откладывают яички в короткостолбчатые женские цветки, то там всегда выводятся личинки и взрослые насекомые; если же яички будут отложены в цветки с длинными столбиками, то, так как яйцеклад бластофаги слишком короток, яички будут отложены в ткань столбика, а не в завязь и погибнут. Но при этом попутно бластофага опылит рыльца длинностолбчатых цветков, и они дадут плоды. Плоды эти, мелкие орешки, остаются внутри вместилища, стенка которого разрастается, становится мясистой, и все оно превращается в "винную ягоду". У большинства разводимых сортов инжира такого опыления и оплодотворения, впрочем, не происходит, и винные ягоды развиваются без него, но семян внутри не содержат (см. партенокарпия, стр. 381).
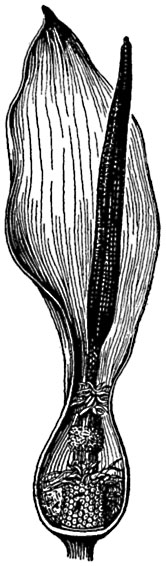
Рис. 310. Соцветие аронника.
Передняя часть покрывала удалена; внизу на початке - женские цветки, над ними - волоски, выше - мужские цветки и над ними - опять волоски. Около женских цветков - мелкие насекомые.
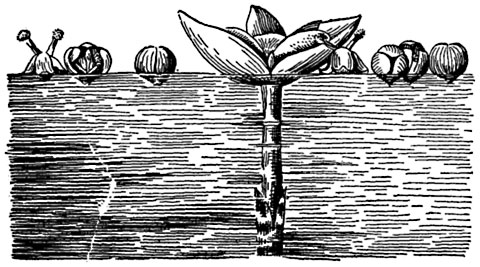
Рис. 311. Опыление у валлиснерии.
В центре - женский цветок на цветоножке, справа и слева от него плавающие мужские цветки на разных стадиях развития.
367
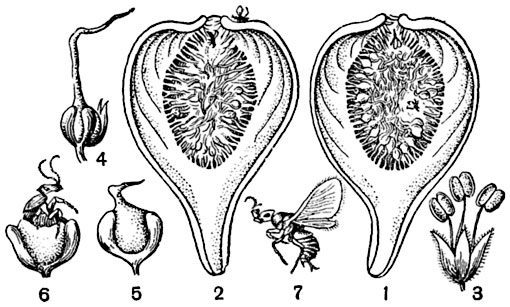
Рис. 312. Перекрестное опыление у инжира:
1 - продольный разрез соцветия, содержащего тычиночные цветки и женские с короткими столбиками, в завязях которых развиваются осы-опылители; 2 - продольный разрез соцветия, содержащего женские цветки с длинными столбиками, в соцветие вползают осы; 3 - тычиночный цветок; 4 - пестичный цветок с длинным столбиком; 5 - пестичный цветок с коротким столбиком; 6 - оса, выходящая из завязи; 7 - оса (увелич.).
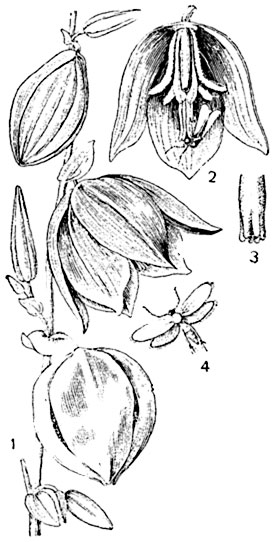
Рис. 313. Опыление у юкки:
1 - ветка из соцветия юкки; 2 - цветок, посещаемый молью, три передние листочка околоцветника удалены; 3 - рыльце пестика; 4 - моль-опылительница.
У некоторых растений опыляющие их насекомые откладывают яички в завязь, где развивающиеся личинки питаются семяпочками, но съедают обычно лишь часть их, а из остальных образуются семена. Это наблюдалось у некоторых видов дрем (Silene nutans, Coronaria flos-cuculi), хлопушек (Silene latifolia), мыльнянки и др.
У американских юкк опыление производится молью пронуба. Она собирает с цветка комочек клейкой пыльцы, летит на другой цветок, откладывает яйцекладом яички в стенку завязи, затем поднимается по столбику на рыльце и запихивает принесенный комочек пыльцы в воронковидное углубление рыльца (рис. 313). Таким образом обеспечивается оплодотворение и дальнейшее развитие семяпочек в семена, причем часть их поедается личинками пронубы. Там, где нет пронуб, юкка обычно не приносит семян.
368
1 Нужно, впрочем, отметить, что цветки некоторых анемофильных растений тоже пахнут, например у клещевины, грецкого ореха, многих пальм, некоторых панданусов и др.
2 Пчелы и шмели, по-видимому, не реагируют на чисто красный цвет.
3 От греческих "дихе" - отдельно, особо, "гамео" - вступаю в брак.
4 От греческих "протерос" - предшествующий, более ранний, "анер" (род. падеж - "андрос") - мужчина.
5 От греческих "протерос" - предшествующий, более ранний,"гюне" - женщина.
6 От греческих "гомос" - вместе, "гамео" - вступаю в брак.
7 От греческих "гетерос" - разный, различный, "стюлос" - столб, колонна, столбик.
8 От греческих "гейтон" - сосед, "гамео" - вступаю в брак.
9 От латинского "легитимус" - законный.
10 От латинского "иллегитимус" - незаконный.
11 Предположения о том, что разные размеры пылинок стоят в связи с тем, что мелкая пыльца при оплодотворении цветков с короткими столбиками развивает более короткие пыльцевые трубочки, не подтвердились: и крупные и мелкие пылинки образуют пыльцевые трубочки одинаковой длины.
12 От латинского "поллен" - очень тонкая мука, пыльца.