

Филогенез листьев
О происхождении листьев существуют различные гипотезы.
В последнее время наибольшим вероятием пользуется теория, основанная на изучении строения первичных сухопутных растений - псилофитов, живших в силурийском и девонском периодах геологической истории Земли.
Впервые они (роды риния, хорнеофитон, астероксилон) были описаны в 1917 г. В настоящее время известно более 20 родов псилофитов,
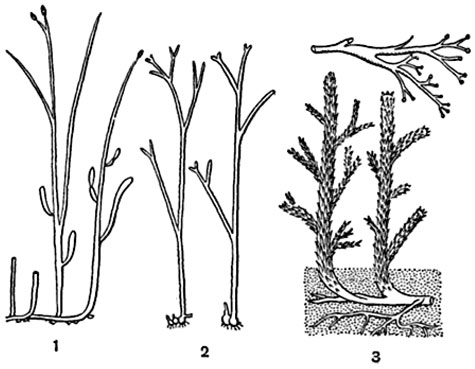
Рис. 241. Псилофитовые.
1 - риния (Rhynia major); 2 - хорнеофитон (Horneophyton); 3 - астероксилон (Asteroxylon); вверху - веточка со спорангиями.
найденных в отложениях различных континентов. У этих растений не было ни корней, ни листьев, и все вегетативное тело их состояло из подземной горизонтальной оси, напоминавшей корневище, и отходящих от нее надземных, дихотомически разветвленных небольших цилиндрических осевых органов, подобных безлистному стеблю и названных теломами1. Концы некоторых разветвлений теломов заканчивались спорангиями (рис. 241). У некоторых псилофитов (астероксилон) теломы были обильно покрыты мелкими щетиновидными или чешуйчатыми выростами. В дальнейшем эволюция этих растений пошла в разных направлениях. У одних маленькие
279
выросты на теломах увеличились в размерах и приняли более или менее плоскую форму, более удобную для фотосинтеза. В них развился проводящий пучок, соединенный с проводящей тканью осевого органа. Это направление эволюции привело к мелколистным высшим споровым растениям - плауновидным и хвощевидным, листья которых, следовательно, развились из мелких выростов на осевых органах, так называемых энациев2. Вторая линия эволюции, приведшая к крупным листьям (как, например, у папоротников), была совершенно иная. Здесь листья возникли из нескольких осевых дихотомических ветвей - теломов - путем перемещения их в одну плоскость, уплощения (изменения радиального строения в дорзовентральное), сращения боковыми частями, утраты способности к неограниченному росту в длину. Листья этого типа по своей морфологической природе подобны кладодиям, имеют "кладодификационное" происхождение (рис. 242).
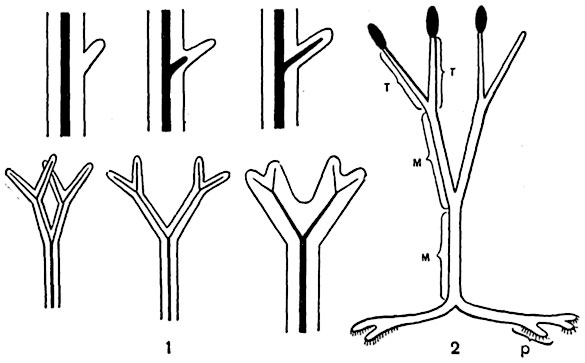
Рис. 242. Схема, иллюстрирующая способ образования листа из теломов псилофитов:
1 - в верхнем ряду путем образования выроста, в нижнем - путем уплощения и срастания теломов; 2 - основные (элементарные) участки тела псилофитов; участки тела, соединяющие теломы (т), носят название месомов (м); подземные участки теломов, давшие затем начало корням, называют ризомоидами (р).
Таким путем развились, вероятно, листья ископаемых и современных папоротников, ископаемых семенных папоротников, саговников, ископаемых беннеттитов и др. Подтверждением этому служит длительный верхушечный рост листьев у папоротников, дихотомическое ветвление жилок у некоторых папоротников и древних голосеменных и некоторые другие признаки крупнолистных растений.
Возникновение у наземных растений листостебельности, т. е. побегов, расчлененных на стебель и листья, было крупнейшим прогрессивным этапом в развитии растительного мира на Земле. Широкие плоские листья значительно увеличили их фотосинтезирующую и транспирирующую поверхность, т. е. значительно увеличили возможность их воздушного и корневого питания (транспирация способствует поглощению и передвижению воды и питательных веществ из почвы).
Если предположить, как это в настоящее время принимается многими ботаниками, что покрытосеменные растения развились из каких-то
280
предков, близких к семенным папоротникам, то листья покрытосеменных возникли из кладодифицированных теломов. Наиболее примитивным типом листьев покрытосеменных многие считают пальчатолопастные листья с пальчатым жилкованием. Из них развились простые, еще менее расчлененные, цельные и цельнокрайние листья. Из пальчатолопастных же, а в ряде случаев, возможно, из простых цельных листьев развились уже вторично сложные и вообще сильно расчлененные листья3. Пальчатое жилкование листьев возникло, по-видимому, из более древнего дихотомического. Из пальчатого жилкования в одном направлении эволюция привела к перистому, характерному для двудольных, а в другом направлении в связи с упрощением листовой пластинки и тоже на ранних стадиях эволюции покрытосеменных - к дуговидному и далее к параллельному, характерным для однодольных.
Прилистники характерны преимущественно для более древних и примитивных двудольных, хотя, впрочем, имеется и ряд исключений из этого положения. Черешок, развивающийся в онтогенезе поздно, когда пластинка уже более или менее сформирована, вероятно, и в филогенезе листьев появился позднее, и примитивные листья предков покрытосеменных были сидячими.
Что касается листорасположения, то одни морфологи считают у покрытосеменных примитивным супротивное листорасположение, другие - очередное (спиральное). Если принять во внимание, что папоротники и примитивные голосеменные, а также более примитивные семейства покрытосеменных имеют очередное листорасположение, то вторая гипотеза имеет больше вероятности, и тогда супротивное и мутовчатое листорасположение нужно считать развившимся из очередного.
281
1 От греческого "телос" - верх, высшая точка. Теломами нередко называют только конечные веточки этих примитивных органов.
2 От латинского "энатус" - произращенный.
3 Это не исключает того, что в ряде родов, видов или рас, расчлененные или малорасчлененные листья могли получиться вторично из сложных, сильно расчлененных, как например у некоторых розоцветных, бобовых и др.