

1. Митоз, или эквационный кариокинез
Непрерывный процесс митоза условно разделяется на следующие фазы: 1) профазу, 2) метафазу, 3) анафазу, 4) телофазу (рис. 20).
1) Профаза. Профаза соответствует процессу формирования определенных, хорошо окрашивающихся структурных единиц, называемых хромосомами (от греческих "хромос" - краска, "сома" - тело).

Рис. 20. Полусхематическое изображение стадий митоза:
1 - интерфаза; 2-4 - профаза; 5-6 - метафаза; 7-9 - анафаза; 10 - переход к телофазе; 11 - телофаза; 12 - поздняя телофаза.
В профазе (рис. 20, 1-4) ядро увеличивается в размерах, хроматиновая сеть становится более грубой, постепенно вместо сети становятся видны нити, вначале тонкие и извилистые. Эти нити создают впечатление рыхлого клубка (отсюда старое название - стадия клубка, или спирема). На самом деле, сплошного клубка не бывает, и формирующиеся хромосомы не соединены в одну запутанную нить. Постепенно нити укорачиваются.
48
уплотняются, окрашиваемость их увеличивается и образуются хромосомы различной величины и формы; они имеют форму палочек, часто дугообразно изогнутых, с равными или неравными плечами; реже они бывают в виде очень коротких палочек и даже почти точек. Уже на ранних стадиях профазы иногда бывает видно, что каждая хромосома расщепляется продольно на параллельно лежащие половинки, называемые хроматидами. Ядрышки, хорошо заметные в начале профазы, постепенно исчезают.
2) Метафаза. Уже в конце профазы на противоположных сторонах ядра в цитоплазме появляется ахроматиновое веретено сначала в виде двух конусообразных участков, отличающихся от цитоплазмы. В живом состоянии эти конусы кажутся бесструктурными, а на фиксированных препаратах в них заметны волокнистые структуры, расходящиеся от вершины. Расположение этих ахроматиновых "колпачков" определяет направление оси деления ядра.

Рис. 21. Развитие сперматозоида хвоща (Equisetum):
1 - последний митоз перед сперматогенезом; 2-4 - формирование сперматозоида, видна центросома и превращение ее в блефаропласт; 5 - сформированный сперматозоид с многочисленными жгутиками.
В животных клетках на концах ахроматиновых конусов наблюдаются еще центросомы, т. е. тельца, от которых во все стороны, как лучи, отходят протоплазматические нити, аналогичные ахроматиновым. В растительных клетках таких центросом большей частью не бывает, но они иногда наблюдаются, особенно при развитии подвижных стадий, там, где они имеются в онтогенезе (у мхов, хвощей и папоротников) при развитии сперматозоидов, а у некоторых растений при образовании зооспор и гамет. В последнем митозе перед их формированием наблюдаются центросомы, превращающиеся затем в опорные базальные тельца, от которых отходят жгутики (рис. 21).
В ранней метафазе (метакинезе) ядерная мембрана исчезает, ахроматиновые нити образуют как бы сплошное веретено. Хромосомы перемещаются в экваториальную часть веретена, образуя так называемую ядерную пластинку (рис. 20, 5, 6). Это стадия поздней метафазы.
Если рассматривать в это время ядерную пластинку в плане, со стороны одного из полюсов веретена, то видно, что каждая хромосома состоит из двух частей - плеч, соединенных более тонким, неокрашивающимся участком - центромерой, или кинетохором. Центромера является точкой перегиба хромосом. Все кинетохоры в метафазе обращены к центру и расположены в одной экваториальной плоскости, где в дальнейшем в большинстве случаев образуется перегородка между новыми клетками. Плечи хромосом иногда могут устанавливаться и в разных плоскостях, но в общем получается звездообразная фигура (отсюда название - стадия звезды, или монастер).
В стадии метафазы хорошо заметна двойственность хромосом. Становится заметно, что каждая хромосома состоит из двух дочерних хромосом, лежащих параллельно друг другу. Это явление иногда называют
49
расщеплением, но это следствие процесса удвоения (редупликации) каждой хромосомы на предыдущих стадиях.
Строение хромосом. Хромосомы существуют в постоянном для каждого вида числе, или, точнее, в двух постоянных числах - гаплоидном и диплоидном, в зависимости от ядерной фазы цикла (см. стр. 300). Гаплоидное число вдвое меньше диплоидного. Так, например, диплоидное число хромосом у сложноцветного Crepis capillaris - 6, столового лука - 16, пшеницы-однозернянки - 14, твердой пшеницы - 28, мягкой пшеницы - 42, хвощей и многих папоротников - более 100, у человека - 46.
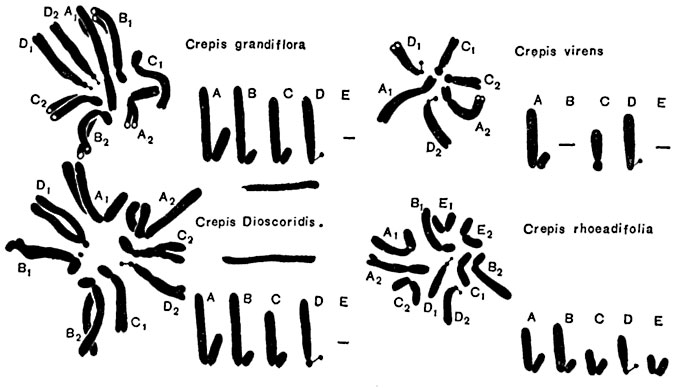
Рис. 22. Набор хромосом у некоторых видов скерды (Crepis). Слева - диплоидная ядерная пластинка (метафаза), справа - гаплоидный набор хромосом.
Размеры хромосом более или менее постоянны и могут служить одним из характерных признаков вида. Длина хромосомы может колебаться от 0,2 до 2 μ. Отдельные хромосомы в ядерной пластинке неодинаковы: крупные и мелкие, длинные и короткие, прямые и согнутые. Плечи хромосом могут быть приблизительно одинаковы (равноплечие) или одно короче другого (неравноплечие), иногда одно плечо настолько коротко, что образует так называемую головку. Некоторые хромосомы имеют так называемых спутников - маленькие хроматиновые тельца, соединенные с хромосомой тонкой нитью. Каждый вид характеризуется определенным набором хромосом, или кариограммой (рис. 22). В диплоидном ядре соответственные по форме и величине хромосомы обнаруживаются в двойном числе и члены каждой пары называются гомологичными хромосомами. Они берут начало от хромосом гаплоидных ядер, слияние которых и дало диплоидный набор хромосом.
Основным веществом в составе хромосом являются ДНК, которые находятся там не только в соединении с белками в виде нуклеопротеидов, но и в свободном состоянии. Поэтому хромосомы проявляют более кислотные свойства, чем протоплазма. Этим в значительной степени объясняется их окрашиваемость основными красками.
Хромосомы обладают более высоким показателем преломления света по сравнению с протоплазмой, благодаря чему они видны в живой делящейся клетке как блестящие тела. Хромосомы обнаруживают двойное луче-
50
преломление, что, по-видимому, объясняется правильной ориентировкой мицелл.
Хромосомы живой клетки нельзя представлять как твердые тела, какими они кажутся на фиксированных и окрашенных препаратах. Вязкость их изменяется в течение митоза; иногда, например, в метафазе, вязкость хромосом особенно велика. С изменениями вязкости, по-видимому, связана пластичность хромосом.
При специальных окрасках хромосом обнаруживается, что каждая хромосома состоит из двух половинок - хроматид. Более тонкие исследования показывают, что хроматида представляет собой спирально скрученную (спирализованную) нить или несколько нитей - хромонем. Спирализация хромосом всегда бывает неравномерной, участки с более плотной спиралью имеют вид зернышек - хромомер. Нередко из-за окраски хромосомы кажутся окруженными гомогенным обволакивающим веществом. Некоторые ученые называли его матриксом (рис. 23). Сейчас его считают артефактом1. В других фазах деления хромосомы менее компактны. Изменение формы хромосом в процессе митоза схематически представлено на рисунке 24. В интерфазе происходит удвоение хромосом (дупликация). Это вполне совпадает с представлениями о воспроизведении ДНК, которое происходит тогда, когда они не закручены в спираль. В профазе можно видеть двойственную природу хромосом. Высказывалось, однако, мнение, что это удвоение начинается уже в метафазе предыдущего деления.
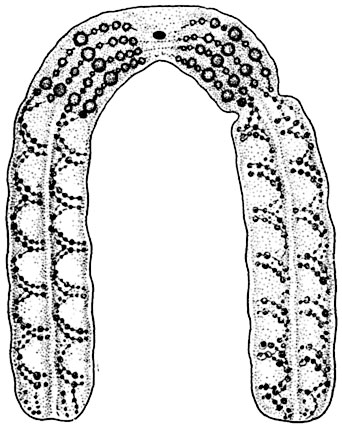
Рис. 23. Схема строения хромосомы.
Видны две хроматиды, одетые матриксом; в каждой хроматиде - по две хромонемы с хромомерами; в месте перегиба - центромера (кинетохор).
Установлено, что хромосомы играют большую роль в явлениях наследственности. Большинство биологов считают, что хромосомы являются основными материальными носителями наследственных свойств организмов; разные хромосомы являются носителями разных наследственных зачатков (генов), которые локализуются в определенных участках хромосом, расположенных линейно, один за другим. С этой точки зрения становится понятным механизм эквационного кариокинеза, при котором расщепление хромосомы по ее длине обеспечивает точное и, следовательно, качественное тождество дочерних хромосом. Наблюдения в световом и электронном микроскопе показали, что хромосомы сохраняют свою индивидуальность и в интеркинетическом ядре. Не следует, конечно, упускать из виду, что хромосомы функционируют в тесной взаимосвязи с цитоплазмой, являясь особо дифференцированными элементами целостной системы всего протопласта. Известны, кроме того, и некоторые случаи так называемой цитоплазматической наследственности, передающейся только через цитоплазму.
51
Аргументами, подтверждающими главную роль хромосом в передаче наследственных свойств, являются следующие:
1) Постоянство числа и формы хромосом, характерное для каждого вида.
2) Механизм митоза, обеспечивающий распределение хромосом между дочерними ядрами.
3) Наблюдаемая нередко корреляция между изменением хромосомы и тем или иным внешним признаком или свойством организма. К этому добавляется еще аналогия между расщеплением свойств в потомстве гибрида и поведением хромосом во время мейоза.
С химической точки зрения молекула ДНК служит важным фактором, объясняющим генетическую непрерывность (наследственная информация).
Указанные факты представляют большой интерес. Из них можно сделать вывод о том, что хромосомы - основной компонент ядра - имеют существенное значение в жизни клетки, но связь их с признаками не непосредственна, а идет через длинную цепь процессов обмена.
С этой точки зрения понятно не только постоянство числа и формы хромосом у различных особей одного вида, но и то, что у родственных видов имеются сходные хромосомы. Как известно, современные систематики нередко с успехом пользуются - для характеристики видов и установления родственных отношений между ними - наряду с внешними признаками также и их кариограммой, или кариотипом, т. е. числом и формой хромосом (рис. 22).
Выше упоминалось, что передача наследственных свойств связана не только с хромосомами. В связи с этим носителем наследственности признается вся клетка в целом.
3) Анафаза. Уже в профазе становится хорошо заметно продольное раскалывание каждой хромосомы на две части, расположенные рядом и параллельно друг другу. В анафазе эти половинки, называемые хроматидами, расходятся к противоположным полюсам веретена (рис. 20, 7-9). Механизм этого расхождения толкуется по-разному. Одни авторы видят его в силах поверхностного натяжения вещества хромосомы, растекающегося от места ее прикрепления вдоль волокон ахроматинового веретена, другие - в сокращении особых тянущих нитей,
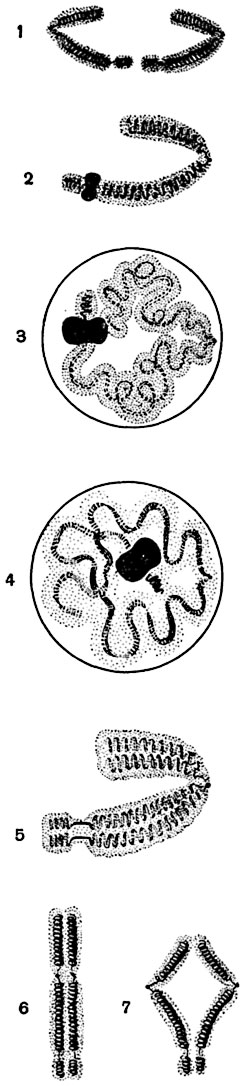
Рис. 24. Схематическое изображение изменения структуры хромосом от поздней анафазы через интеркинез и профазу до следующей анафазы:
1 - поздняя анафаза; 2 - телофаза; 3 - интерфаза; 4 - интерфаза, переход к профазе; 5 - поздняя профаза, переход к метафазе; 6 - метафаза; 7 - анафаза.
52
идущих от полюса веретена и прикрепляющихся к кинетохорам хромосом. Путь, проделываемый хромосомами, относительно размеров клетки довольно велик (от 5 до 25 μ). Скорость перемещения равна примерно 1 μ в минуту.
4) Телофаза представляет процесс реконструкции двух дочерних ядер из разошедшихся к противоположным полюсам ахроматинового веретена двух групп хроматид. В общем этот процесс представляет как бы зеркальное отображение профазы, но идущей в обратном направлении (рис. 20, 10 - 12).
Хромосомы утоньшаются, удлиняются, теряют мало-помалу способность окрашиваться; получается структура интеркинетического ядра; появляются ядерная оболочка, кариолимфа и ядрышки.
В анафазе на фиксированных и окрашенных препаратах между двумя группами разошедшихся хромосом выявляются нити, похожие на нити ахроматинового веретена, но имеющие, возможно, независимое от него происхождение. В телофазе эти так называемые соединительные нити делаются обильней, общий пучок их, расширяясь посередине, принимает форму бочонка - фрагмопласта, достигающего своими выпуклыми сторонами боковых стенок клетки. Затем на этих нитях в экваториальной плоскости появляются уплотнения, которые сливаются потом в срединную пластинку (мембрану), перегораживающую общую полость клетки на две - каждую с одним ядром. Таким образом, вслед за митозом происходит деление клетки (рис. 20).
Абсолютная и относительная продолжительность фаз митоза различна у разных объектов и сильно зависит от температуры и других внешних факторов. Продолжительность всего процесса митоза в тычиночных волосках традесканции при 10-11°С - 135 минут, при 25°С - 75 минут, а при 45° С - только 30 минут. Существенна продолжительность отдельных фаз. У той же традесканции продолжительность профазы была 103 минуты, метафазы - 14 минут, анафазы - 15 минут, телофазы - 8 минут. Последнее число, несомненно, занижено, так как телофаза учтена здесь только до момента образования оболочки, хотя реконструкция ядра продолжается и после этого. В общем можно сказать, что профаза и телофаза наиболее длительны, занимая вместе более
| 3 |
| 4 |
Образование перегородок между клетками и образование новых клеток. Вслед за делением ядра происходит деление протопласта. В конце деления ядра образуется перегородка, общая обеим дочерним клеткам: - мембрана, или срединная пластинка. Срединная пластинка большей частью возникает начиная от центра клетки и распространяется к стенкам. Эта перегородка является, вероятно, не сплошной, а пронизана тончайшими сквозными ходами ("плазмодесменными канальцами"), в которых тянутся из одной клетки в другую, смежную, тонкие нити плазмы - плазмодесмы, связывающие соседние протопласты друг с другом (стр. 97). В дальнейшем в результате деятельности цитоплазмы на срединную пластинку накладываются оболочки каждой из клеток.
В ряде случаев образование оболочки клеток происходит несколько иначе.
53
В некоторых многоядерных быстрорастущих клетках, например в зародышевом мешке, при развитии эндосперма проходит сначала целый ряд митозов и только потом образуются перегородки клеток (рис. 56). У некоторых растений образование дочерних клеток происходит не в результате появления перегородок между дочерними ядрами, а в результате процесса, называемого почкованием.
При почковании материнская клетка образует округлый или эллипсоидальный вырост, постепенно увеличивающийся. Ядро материнской клетки делится, и одно из дочерних ядер перемещается в вырост. Путем суживания перетяжки между клеткой и ее выростом происходит отшнуровывание выроста - образуется дочерняя клетка. Отпочковавшаяся клетка может, еще не вполне отделившись от материнской, в свою очередь почковаться. У дрожжевых грибов при определенных условиях путем почкования образуются ветвистые цепочки клеток.
54
1 Артефакт - посмертное изменение, не существующее в живой клетке.