
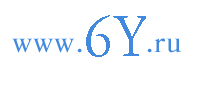

ЯДРО
Строение ядра
Клеточное ядро (nucleus) представляет обязательную часть содержимого живой клетки как животных, так и растений.
Впервые ядро было открыто в 1831 г. в клетках орхидей Робертом Броуном, а затем замечено и в клетках всех других растений. Особенно большое внимание стали уделять изучению строения ядра в 80-х годах прошлого столетия, когда было установлено, что деление ядра предшествует делению клетки1.
Ядро всегда полностью погружено в протоплазму и никогда не переходит в вакуолю, как это часто бывает, например, с крахмальными зернами.
В большинстве случаев каждая клетка содержит по одному ядру, но у
40
некоторых, особенно у низших, растений распространены и даже преобладают многоядерные клетки.
Форма и величина клеточных ядер различны у разных растений и в разных органах их.
Большей частью ядра имеют округлую или чечевицеобразную форму. Нередко наблюдается известное соотношение между формой клетки и содержащегося в ней ядра: в изодиаметрических клетках ядра обычно круглые, а в вытянутых - веретеновидные и даже нитевидные, вытянутые вдоль оси клетки (рис. 13).
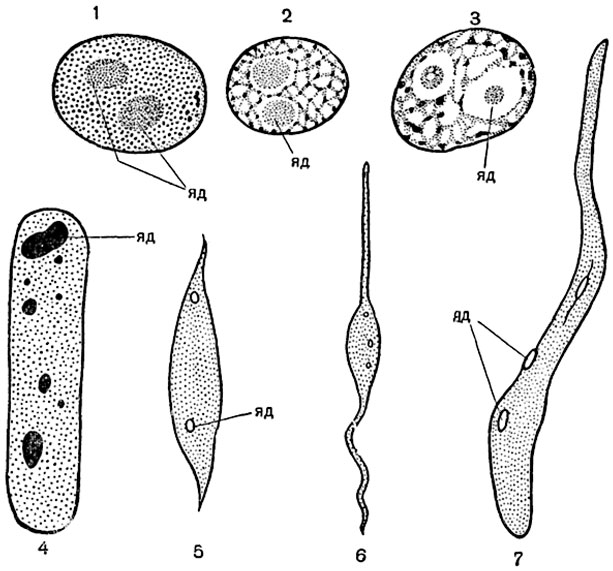
Рис. 13. Формы ядра:
1, 2, 3 - ядра из клеток корня гиацинта (Hyacinthus); 4 - ядро из вытянутой в длину клетки птицемлечника (Ornithogalum); 5, 6 - ядро с жгутиковидньши образованиями из клеток черешка листа пеларгонии (Pelargonium); 7 - ядро из слизевых клеток алоэ (Aloe); яд - ядрышки.
В молодых клетках ядро занимает относительно больший объем протопласта, чем в клетках сформированных, закончивших свой рост. В молодых клетках отношение объема ядра к объему протопласта составляет от
| 1 |
| 4 |
| 1 |
| 6 |
| 1 |
| 20 |
| 1 |
| 200 |
У высших растений ядра средних размеров имеют диаметры 10-20 μ. Исключительно крупные круглые ядра встречаются в яйцеклетках саговников, где они достигают 600 μ в диаметре, т. е. ясно различимы невооруженным глазом.
Строение ядра различно в состоянии деления и между делениями. Последнее называется часто покоящимся состоянием, хотя это по существу
41
неверно, так как в этот период в ядре наиболее активно происходят процессы обмена веществ. Лучше называть такое "покоящееся" ядро интеркинетическим, т.е. находящимся в промежутках между делениями. В таком состоянии ядра чаще всего и приходится наблюдать. Поэтому с него мы и начнем наше описание.
В живых, совершенно неповрежденных клетках высших растений ядра нередко мало отличаются по светопреломлению от окружающей цитоплазмы, так что при наблюдении слабо различимы в виде пузырька со светлым контуром (рис. 14). В таком пузырьке резко выступают одно или несколько ядрышек в виде круглых, сильнее преломляющих свет (блестящих) телец. После фиксации и окраски ядра делаются видимы значительно яснее.

Рис. 14. Живые клетки эпидермиса чешуи лука при наблюдении в темном поле:
А - ядро совершенно неразличимо; Б - ядро различимо в виде светлого пузырька (легкое повреждение клетки).
Фиксация заключается в обработке живых клеток ядовитыми для них веществами, так называемыми фиксаторами, такими, как спирт, формалин, сулема, хромовая кислота, уксусная кислота и др., употребляющимися как в отдельности, так и в смеси друг с другом. Действие их выражается в основном в необратимой коагуляции белковых и иных коллоидов клеточного содержимого, и получившиеся сгустки, имеющие более сильную светопреломляемость, делаются яснее различимыми под микроскопом в виде тех или иных структур. Они делаются еще яснее после окраски, избирательно окрашивающей отдельные структуры. Для ядра особое значение имеют основные, или так называемые ядерные, краски, такие, как гематоксилин, фуксин, метиленовый синий и др., ярко окрашивающие так называемые хроматиновые структуры его (от греческого "хрома" - окраска). Ввиду указанных преимуществ в четкости получаемых картин при такой обработке строение клеточных ядер и изучалось больше на окрашенных препаратах, чем в живом состоянии. При этом методика фиксаций и окрасок нередко воспринималась некритически, без учета того, насколько она передает истинные структуры, существовавшие в живом ядре, и насколько дает артефакты, т. е. создает новообразования в результате коагулирующего действия фиксатора.
Ядро фиксированной клетки, находящееся в интеркинетическом состоянии, содержит ядерный сок, или кариолимфу, в котором расположены тонкая хроматиновая нитчато-зернистая структура - хроматин и одно или несколько ядрышек, особенно ярко окрашивающихся основными красителями благодаря высокой концентрации рибонуклеопротеидов. От цитоплазмы ядро отграничено очень тонкой цитоплазматической мембраной (ядерной оболочкой). Электронный микроскоп позволяет видеть, что мембрана двуслойна и в ней имеются очень тонкие пронизывающие ее канальцы (рис. 17). Оболочка ядра представляет собой часть мембранной системы клетки. Между ядром и цитоплазмой осуществляется постоянный обмен веществ. Описанные структуры хорошо видны на фиксированных и окрашенных препаратах. Наличие описанных структур подтверждают и фотографические снимки с живых, нефиксированных и неокрашенных ядер.
Существовали взгляды, согласно которым в живом интеркинетическом ядре, кроме ядрышек, нет никакой структуры, и вся полость ядра заполнена жидкой гомогенной массой, по своей вязкости близкой к глицерину. Однако из сказанного выше понятно, что оптическая гомогенность живого ядра не означает его структурной гомогенности.
42
По химическому составу и физико-химическому строению как ядро, так и цитоплазма представляют в основном комплекс гидрофильных коллоидов, находящихся в большей или меньшей степени набухания. Из них главное значение имеют белки, и особенно сложные белки, называемые протеидами (стр. 32), представляющие соединения собственно белка (протеина) с небелковым компонентом, большей частью кислотного характера. Из них особенно важное значение имеют нуклеопротеиды - соединения белка с нуклеиновыми кислотами .
Известны два типа нуклеиновых кислот: содержащиеся в ядре дезоксирибонуклеиновые (тимонуклеиновые), сокращенно называемые ДНК, и содержащиеся в ядрышках и цитоплазме рибонуклеиновые кислоты, обозначаемые РНК. Они обусловливают окрашиваемость ядра основными красками, так как лежат в основе хроматина2. ДНК весьма точно обнаруживаются цветной реакцией Фельгена3. РНК этой реакции не дают. ДНК входят в состав клеточного ядра в виде нуклеопротеидов, а отчасти, вероятно, и в свободном состоянии.
Нуклеиновые кислоты представляют собой очень длинные цепочки, звенья которых - нуклеотиды - состоят из пятиуглеродного сахара (рибозы C5H10O5 или дезоксирибозы C5H10O4), фосфорной кислоты и одного из азотистых оснований.
Макромолекула нуклеиновой кислоты состоит из двух таких цепей, обвивающих одна другую и образующих таким образом двойную спираль (рис. 15). Расположение нуклеотидов в молекуле можно представить себе следующим образом:
| фосфат фосфат |
╲ ╱ |
сахар |
— |
основание |
—— |
основание |
— |
сахар |
╱ ╲ |
фосфат фосфат |
| фосфат фосфат |
╲ ╱ |
сахар |
— |
основание |
—— |
основание |
— |
сахар |
╱ ╲ |
фосфат фосфат |
| фосфат фосфат |
╲ ╱ |
сахар |
— |
основание |
—— |
основание |
— |
сахар |
╱ ╲ |
фосфат фосфат |
| фосфат фосфат |
╲ ╱ |
сахар |
— |
основание |
—— |
основание |
— |
сахар |
╱ ╲ |
фосфат фосфат |
| 3 | 2 | 1 | 2 | 3 |
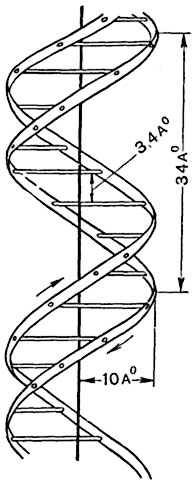
Рис. 15. Схема строения молекулы ДНК.
Каждая лента представляет собой основную цепь. Горизонтальные отрезки соответствуют парам оснований, присоединенных по одному к каждой цепи.
43
В схеме показано расположение водородных (1), глюкозидных (2) и сложноэфирных (3) связей.
Основания, входящие в состав РНК и ДНК, несколько различны. ДНК содержат пиримидиновое основание тимин (Т), а РНК - урацил (У). Пиримидиновое основание цитозин (Ц) и пуриновые основания аденин (А) и гуанин (Г) входят в состав обеих кислот.
В любом участке молекулы основание одной цепи располагается против основания другой.
Соединяются основания только в определенных сочетаниях, именно А и Т, Г и Ц; в цепи располагаться они могут в любом порядке, но только указанными парами. Эти цепи, соединенные основаниями и дополняющие друг друга (комплиментарные), несколько напоминают негатив и позитив. ДНК обладают способностью синтеза подобных себе ДНК. В определенных условиях цепи разделяются вдоль и каждая из них (половина молекулы) строит недостающую (рис. 16). В результате формируются две совершенно идентичные молекулы. ДНК определяет синтез соответствующих РНК (рис. 16, 17). Ряд экспериментов позволил установить, что по ДНК строятся РНК, которые в свою очередь обеспечивают установление определенной последовательности аминокислот в белках (рис. 17). ДНК участвуют в синтезе ферментов. Для осуществления этих процессов необходимы определенные ферменты, энергия и исходные материалы.
Среди растений только сине-зеленые водоросли и многие бактерии не имеют типично оформленных ядер. Однако у тех и других в протоплазме
Рис. 16. Схематическое изображение процесса, при помощи которого происходит воспроизведение ДНК и построение ею РНК, направляющей в свою очередь синтез белков.
Четыре типа нуклеотидов ДНК изображены в виде попарно дополняющих друг друга строительных блоков: А - аденин; Г - гуанин; Т - тимин; Ц - цитозин. Одна цепь ДНК (показана косой штриховкой) строит подле себя из нуклеотидов новую цепь либо ДНК (I), либо РКН (II)(показано штриховкой клеткой); эта новая полинуклеотидная цепь комплиментарна исходной цепи.
44
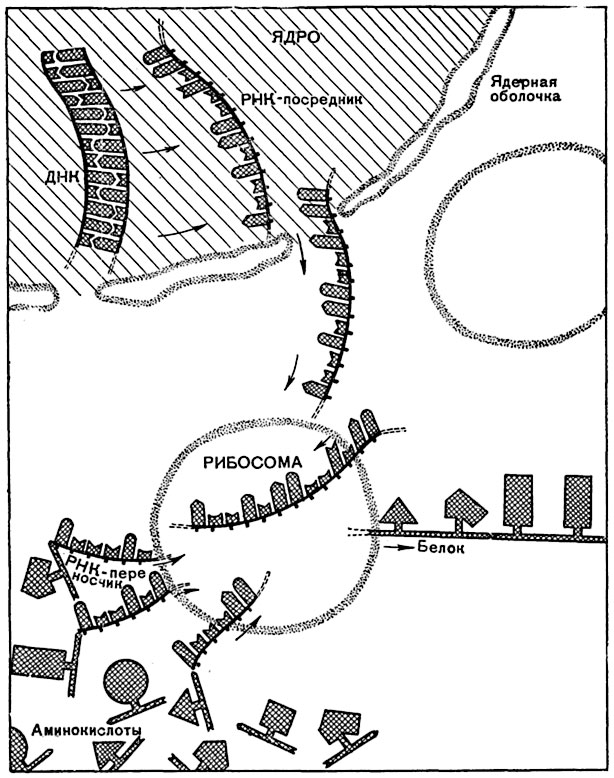
Рис. 17. Схема синтеза белка в клетке, в котором участвуют ДНК, РНК-посредник, РНК-переносчик и химические группировки, богатые энергией.
Молекула РНК-переносчика содержит примерно 80 оснований, а молекула РНК-посредника - 500 или более.
содержится большой процент нуклеопротеидов, входящих в состав ядер у остальных растений. Далее, не содержат ядер клетки ситовидных трубок высших растений, но это дегенеративное явление: в начале развития каждая такая клетка имеет как ядро, так и протоплазму, но впоследствии первое дегенерирует, а вторая остается.
45
1 И. Д. Чистяков наблюдал деление ядра при образовании спор у хвощей (1874).
2 Нельзя отождествлять дезоксирибонуклеиновую кислоту и хроматин: первая есть химическое понятие, а второй - морфологическое, притом неоднородное в химическом отношении, хотя в состав его всегда входит дезоксирибонуклеиновая кислота.
3 Проведение реакции Фельгена сравнительно сложно. Ограничимся небольшим пояснением. Реакция заключается в том, что срезы гидролизуют однонормальной соляной кислотой 8 - 10 минут в термостате при 60°С и затем помещают в раствор фуксинсернистой кислоты (полученный в результате действия на раствор основного фуксина сначала соляной кислоты, а затем бисульфита натрия NaHSO3). ДНК с этим раствором дают розово-фиолетовое окрашивание. Возможно приготовление постоянных препаратов.