
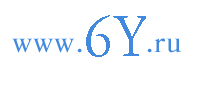

ПРОТОПЛАСТ
Протопласт1 состоит из протоплазмы (называемой иначе цитоплазмой2 или, короче, плазмой), одного или нескольких клеточных ядер, пластид, хондриосом, митохондрий, рибосом, лизосом и др. (рис. 1, 2).
Цитоплазму и находящиеся в ней ядра, пластиды, хондриосомы, рибосомы, аппарат Гольджи и др. называют органеллами или органоидами клетки.
В протопласте обычно находятся вакуоли - пространства, заполненные водой с растворенными в ней минеральными и органическими веществами, представляющими продукты обмена веществ (рис. 2, 3, 4). Многие из этих веществ встречаются и в твердом виде как в цитоплазме, так и в органеллах клетки.
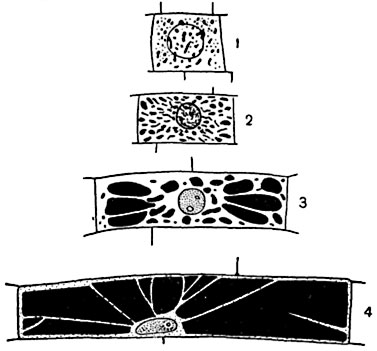
Рис. 3. Формирование вакуолей в клетке эпиблемы корня пшеницы (Triticum vulgare), изображенное по препаратам с прижизненной окраской нейтральной красной.
Клетки: 1 - из верхушечной меристемы; 2 - из основания зоны растяжения; 3 - из середины зоны растяжения; 4 - из зоны всасывания.
Снаружи протопласт покрыт твердой оболочкой; у некоторых клеток она отсутствует в течение всей жизни клетки или определенных стадий ее развития. Иногда встречаются клетки, не имеющие ядер, либо не имеющие ядер, пластид и хондриосом.
ЦИТОПЛАЗМА (ПРОТОПЛАЗМА)
На протоплазму простейших (Protozoa) в 1835 г. обратил внимание французский биолог Феликс Дюжарден, назвавший ее саркодой. Термин "протоплазма", как наименование живого вещества (за исключением ядра) яиц и клеток зародыша животных, ввел в биологию в 1840 г. чешский анатом и гистолог Ян Пуркыня.
Протоплазма бесцветна, прозрачна. Оптически, даже при рассматривании в ультрамикроскоп, протоплазма представляется почти пустой. Она сильнее, чем вода, преломляет свет. Частицы ее анизотропны, обладают двойным лучепреломлением.
Удельный вес протоплазмы был определен 1,025-1,055, в отдельных случаях найден несколько более низким (1,010) или более высоким (1,060).
Консистенция протоплазмы варьирует от водянистого золя, как во
29
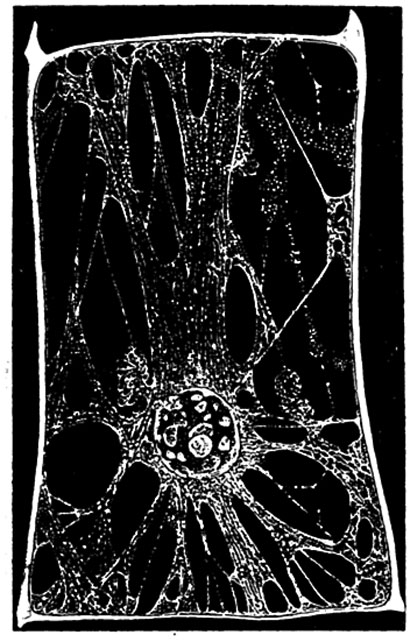
Рис. 4. Клетка из волоска стебля тыквы (Cucurbita pepo), увеличение около 120 раз. Вакуоля пронизана многочисленными тяжами протоплазмы, соединяющими ядерный кармашек с постенным слоем. Видны многочисленные органеллы и включения.
многих деятельных клетках, до тягуче-вязкого, плотностуденистого и даже до твердого геля (студня), как в сухих покоящихся спорах и семенах.
Цитоплазма состоит из трех слоев. Самый наружный слой - плазмалемма - прозрачен и не содержит никаких органелл. Вероятнее всего, его мицеллы правильно ориентированы. Средний слой - мезоплазма - наиболее мощный, кажется ячеистым и содержит все оргалеллы. Третий слой - тонопласт - окружает вакуоли и так же, как плазмалемма, состоит из правильно ориентированных мицелл, но значительно более мощный (рис. 5). Во многих исследованиях цитоплазма разделяется на два слоя: эктоплазму и эндоплазму, в которой и содержатся все органеллы.
Исследования при помощи электронного микроскопа показали, что цитоплазма состоит из основной цитоплазмы - гиалоплазмы3 и расположенной в ней системы двойных мембран4 - эндоплазматической сети, или ретикулюма. Между мембранами находятся вещества, составляющие активный компонент цитоплазмы. Элементы ретикулюма изменяются в зависимости от физиологического состояния клетки (рис. 6). Цитоплазма представляет собой материю, обладающую формой движения, называемой жизнью. Ей свойственны все явления, которые типичны для понятия "жизнь", - питание, дыхание, рост, движение.
Химический состав протоплазмы. Протоплазма представляет чрезвычайно сложный комплекс веществ, с более или менее значительным содержанием воды; этот комплекс имеет изменчивую (лабильную) физико-химическую структуру.
Соединения, входящие в состав протоплазмы, очень многочисленны; их разнообразие и высокая реакционная способность создают в клетке весьма большие возможности для осуществления самых различных
30
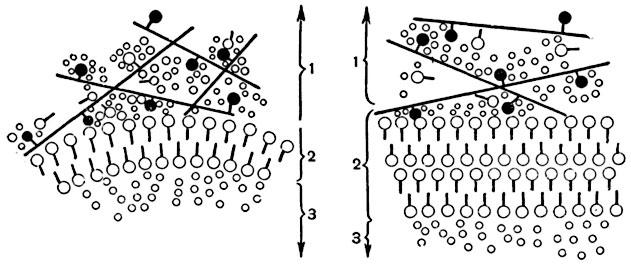
Рис. 5. Схема субмикроскопического строения тонопласта:
1 - мезоплазма; 2 - тонопласт; 3 - вакуоля. Черными и белыми кружками с черточками и линиями обозначены коллоиды белкового происхождения. В тонопласте липопротеиды ориентированы определенным образом, в мезоплазме этой правильности не заметно.
процессов. В живой клетке эти процессы строго согласованы, и эта согласованность обусловливает нормальную жизнедеятельность клетки.
Химический анализ протоплазмы в лаборатории лишь с первой степенью приближения выясняет ее состав: уже при подготовке к анализу нарушается естественная структура протоплазмы, ферментативным или даже чисто химическим путем могут образоваться новые вещества или, наоборот, разрушиться существующие в живой протоплазме лабильные (неустойчивые) соединения.
Весьма нелегка даже в первом приближении задача определения химического состава протоплазмы многоклеточных растений с их мощной системой клеточных стенок и одноклеточных растений, снабженных твердой оболочкой; отделить протопласты от их оболочек и выделить из цитоплазмы другие органеллы, содержимое вакуолей и т. п. технически трудно.
Для химического анализа протоплазмы сравнительно удобным объектом являются слизевики (иначе, слизистые грибы, или миксомицеты). В определенной стадии онтогенеза слизевики представляют плазмодии - крупные тела, состоящие почти нацело из цитоплазмы и ядер, если не считать частиц перевариваемой пищи (преимущественно в виде микроорганизмов) и продуктов запаса и отброса.
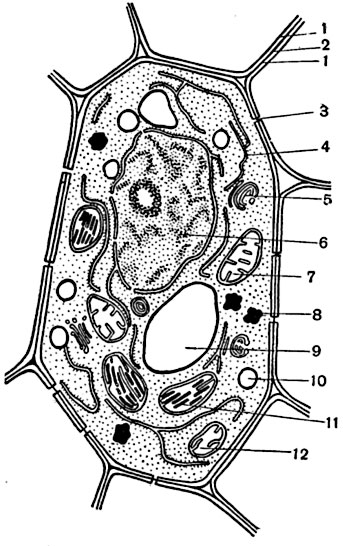
Рис. 6. Схема строения живой клетки растения:
1 - первичная стенка; 2 - срединная пластинка; 3 - плазмодесмы; 4 - эндоплазматический ретикулюм; 5 - аппарат Гольджи; 6 - ядро с ядрышком; 7 - митохондрии; 8 - капли жира; 9 - вакуоля; 10 - крахмальные зерна; 11 - хлоропласты; 12 - пропластиды.
31
Анализы миксомицета Reticularia lycoperdon дали следующие результаты (в процентах от сухого вещества плазмодиев):
| Белок (кроме пластина) вместе с нуклеопротеидом . . . . . . . . . . . . . . . . . . . . . . . . . . . . | 20,65 |
| Пластин (альбуминоподобный белок) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . | 8,42 |
| Нуклеиновая кислота (свободная и связанная) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . | 3,68 |
| Азотистые экстрактивные вещества . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . | 12,00 |
| Масло вместе с пигментом . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . | 17,85 |
| Лецитины . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . | 4,67 |
| Холестерины . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . | 0,58 |
| Масло липопротеидов . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . | 1,20 |
| Летучие кислоты . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . | + |
| Восстанавливающие углеводы . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . | 2,74 |
| Невосстанавливающие углеводы (без гликогена, трегалоза) . . . . . . . . . . . . . . . . . . . . . | 5,32 |
| Гликоген . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . | 15,24 |
| Микоглюкозан5 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . | 1,78 |
| Неизвестные вещества . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . | 5,87 |
Главнейшими не только по количеству, но и по значению составными частями протоплазмы являются белковые вещества (белки); они составляют (см. таблицу) значительный процент сухого вещества протоплазмы, и они же представляют собой важнейшую материальную основу жизненных явлений.
В самом деле, по определению Фридриха Энгельса, "жизнь есть способ существования белковых тел, существенным моментом которого является постоянный обмен веществ с окружающей их внешней природой, причем с прекращением этого обмена веществ прекращается и жизнь, что приводит к разложению белка"6.
Из элементов белковые вещества содержат углерод (в количестве 55- 56%), кислород (20 - 25%), азот (15-19%), водород (6,5-7,5%); некоторые белковые вещества содержат еще серу (0,3-2,5%) и фосфор.
Молекулы белковых веществ очень крупны и имеют весьма сложное строение.
В задачу курса не входит подробное описание белков, достаточно указать, что существуют простые и сложные белки.
Простые белки, собственно белки, или протеины, при гидролизе распадаются только на аминокислоты.
Аминокислоты получаются из ряда органических кислот, в которых атом водорода заменяется аминогруппой (NH2), отсюда и происходит их название. Для них дается формула R · CHNH2 · COOH. Радикал R принадлежит к жирному или к ароматическому ряду или включает и гетероциклические кольца. Содержащиеся в клетках свободные аминокислоты представляют собой продукты как синтеза, происходящего внутри клеток, так и непрерывного расщепления ранее сформированных белков.
Аминокислоты могут вступать в соединение друг с другом с выделением воды, образуя длинные молекулы так называемых пептидов. Молекулы пептидов могут состоять из остатков молекул различных аминокислот или одной и той же аминокислоты. Число аминокислотных остатков, составляющих белки, колеблется от нескольких сотен до нескольких тысяч.
К простым белкам, притом сравнительно легко выделяемым из содержимого клеток растений, относятся запасные белковые вещества семян.
В состав сложных белков, или протеидов, кроме простых белков, входят вещества небелкового происхождения - простетические группы. К этим белкам относятся нуклеопротеиды, которые играют важную роль в жизни клетки, их простетические группы представляют собой нуклеиновые кислоты; гликопротеиды, у которых белок связан с углеводом; липопротеиды, представляющие сочетание простых белков с жирными кислотами; хромопротеиды - биологически очень важные вещества, в которых протеин соединен с пигментом.
Протеиды представляют собой существенную составную часть протоплазмы и клеточных ядер.
Наиболее показательны следующие реакции для обнаружения белков под микроскопом:
32
1. Ксантопротеиновая реакция. Под действием крепкой азотной кислоты белки окрашиваются (особенно при подогревании) в желтый цвет; если прилить раствор аммиака, окраска переходит в оранжевую; если прибавить вместо аммиака едкого натра (в избытке), то получается красновато-коричневое окрашивание. Под микроскопом окраска обычно представляется слабой.
2. Миллонова реакция. В растворе азотнокислой ртути, приготовленном определенным образом и содержащем окислы азота, белки дают красное окрашивание (розового, малинового или кирпичного оттенка); окраска более интенсивна при подогревании препарата с раствором до кипения.
3. Биуретовая реакция. При действии на белок раствора едкой щелочи, с прибавлением небольшого количества (1-2 капель) слабого (например , 2%-ного) раствора медного купороса (CuSO4) появляется фиолетовое окрашивание; если прибавить еще CuSO4, окраска переходит в синеватую.
4. Реакция Распайля. При действии концентрированного раствора сахара и концентрированной серной кислоты белки дают красное окрашивание различных оттенков (чисто красное, пурпуровое, фиолетово-красное); под микроскопом окраска кажется розовой.
Указанные реакции есть в сущности реакции на те или иные продукты распада белка. Чтобы твердо установить наличие или отсутствие белков в препарате, необходимо применить и сопоставить несколько реакций и дополнить исследование пробой на окрашивание йодом, анилиновыми красками.
Йод применяется в 5-10%-ном растворе в спирту или в растворе с добавлением йодистого калия; он окрашивает белковые вещества в желтый или коричневый цвет. Из анилиновых красок употребляются - в водном или в спиртовом растворе - фуксин, гематоксилин. Йод и названные краски вызывают коагуляцию (свертывание) белков и отмирание протоплазмы. Некоторые анилиновые краски употребляются для прижизненного окрашивания протоплазмы; под действием очень слабых растворов эозина, метилвиолета, хризоидина протоплазма, окрашиваясь, остается в течение некоторого времени живой.

Рис. 7. Схема частицы (мицеллы) гидрофильного коллоида.
Кроме белков и аминокислот протоплазма содержит минеральные соли, углеводы, жиры, липоиды и целый ряд других веществ. Исследования с помощью радиоактивных изотопов показали, что в клетках происходит постоянное обновление химического состава протоплазмы. Физико-химическое состояние ее также весьма изменчиво.
Коллоидное состояние. Со стороны физико-химического состояния протоплазму можно охарактеризовать как вязкую сложную коллоидную7 систему. Коллоиды протоплазмы относятся к так называемым лиофильным (гидрофильным) коллоидам.
Коллоидная система в данном случае состоит из воды (дисперсионная среда), в которой находятся в мелкораздробленном состоянии различные вещества в виде частиц размерами от 0,001 до 0,1 μ (дисперсная фаза). Частицы дисперсной фазы состоят обычно из нескольких молекул. Некоторые высокомолекулярные органические вещества, как белки, полисахариды (например, крахмал), имеют размеры молекул порядка нескольких мμ и образуют вместе с дисперсионной средой коллоидные системы даже в молекулярном состоянии.
Частицы дисперсной фазы, находясь в дисперсионной среде, имеют громадную поверхность соприкосновения со средой и, следовательно, огромные возможности для различных химических и физических процессов. В коллоидных растворах частицы дисперсной фазы оказываются прочно связанными непосредственно или посредством адсорбционного слоя третьего вещества - стабилизатора - с молекулами дисперсионной среды, образующей вокруг частиц так называемые сольватные или, если дисперсионной средой является вода, гидратные оболочки (рис. 7). Сольватированные частицы носят название мицелл. Часто концы их несут электрические заряды. При потере мицеллами сольватных оболочек и зарядов происходит коагуляция (свертывание); коллоидные частицы слипаются, диспергированная фаза
33
выпадает в осадок. При неполной коагуляции, называемой коацервацией8, коллоидный раствор (золь) расслаивается на коацерват, богатый коллоиднодиспергированным веществом, и равновесную жидкость, бедную диспергированным веществом или вовсе его не содержащую.
При коацервации частицы диспергированной фазы теряют диффузную оболочку (б), сохраняя сольватную (а) (рис. 8). При сближении таких частиц их сольватные оболочки объединяются, и частицы сливаются друг с другом (рис. 8). Между коацерватом и равновесной жидкостью имеется поверхность раздела. Некоторые ученые считают протоплазму сложной системой коацерватов в равновесной жидкости - воде.
Некоторые ученые (А. И. Опарин, Бунгенберг де Йонг) считают, что протоплазма представляет собой систему коацерватов, которая содержит почти нацело связанную воду; связанная вода обладает пониженной растворяющей способностью и обеспечивает несмешиваемость протоплазмы с обычной водной средой; в силу этого протоплазма ведет себя как особая фаза.
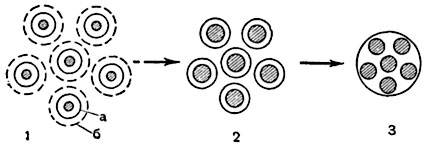
Рис. 8. Схема образования коацервата (1-3):
а - плотный слой сольватной оболочки; б - диффузная оболочка.
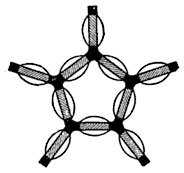
Рис. 9. Схема структуры геля.
Коллоиды протоплазмы обладают способностью изменять свое состояние и переходить из золя в гель (студень). Коагуляции с разделением на твердую и жидкую фазы при этом не происходит.
По-видимому, можно предположить, что при этом процессе мицеллы, имеющие форму палочек, одеты гидратными оболочками только в средней части; оба конца мицеллы несут электрические заряды. Такие мицеллы не соединяются друг с другом, и коллоид находится в состоянии золя.
При утрате зарядов частицы соединяются концами (рис. 9) и образуется сетчатый остов с жидкостью в ячеях сетки. В результате получается структура с непрерывной твердой и с непрерывной жидкой фазой; это и есть гель. В некоторых случаях при образовании геля цепочкообразные мицеллы переплетаются, образуя сеть. В ячейках этой сети удерживается вода.
При 50-60°С белковые вещества протоплазмы обычно претерпевают необратимую коагуляцию и протоплазма (а с нею клетка и весь организм) отмирает. Однако некоторые растения (из семейств кактусов, толстянковых) выносят без вреда нагревание солнцем до 50-60° С, некоторые бактерии и сине-зеленые водоросли живут и размножаются в горячих источниках с температурой около 70° С и выше. В сухом состоянии (в семенах, в спорах низших растений) протоплазма сохраняет жизнеспособность и после воздействия, в течение некоторого времени, температуры в 100° С.
Вязкость протоплазмы. Вязкость характеризует состояние коллоидов протоплазмы и вместе с тем отражает физиологическое состояние не только клетки и органа, но и всего организма. Вязкость зависит от температуры, pH и других условий.
Протоплазма не смешивается с водой и не расплывается, как должна была бы в ней расплываться, если бы представляла собой обычный гидрозоль.
Вязкость протоплазмы различна у разных видов растений. Она бывает в 12-20 раз выше вязкости воды (в 80-100 раз меньше вязкости касторового масла). В клетках стебля кормовых бобов протоплазма оказалась в 24 раза более вязкой, чем вода, и в 13 раз более вязкой, чем клеточный сок
34
этих же клеток. У водных растений она значительно меньше, только в 5-6 раз превышает вязкость воды.
В клетках вегетативных органов молодых растений вязкость протоплазмы невелика, по мере роста растений постепенно увеличивается и затем, к цветению, уменьшается; по отцветании увеличивается вновь. В клетках цветка вязкость очень велика.
Эластичность протоплазмы9. Живая протоплазма очень эластична. При помощи микроманипулятора10 протоплазму можно растянуть в очень тонкую нить. Если ее отпустить, она снова примет прежнюю округлую форму.
Эластичность протоплазмы хорошо демонстрируется в опытах с плазмолизом (стр. 68). Эластичность протоплазмы, так же как и вязкость, зависит от состояния растения.
Полупроницаемость. Одним из очень важных свойств протоплазмы живой клетки является ограниченная избирательная проницаемость, или полупроницаемость. Процесс питания, снабжения клеток водой, накопления запасов связаны с передвижением веществ из одних клеток в другие, поэтому важную роль в жизни клеток и всего организма играет поступление воды и растворенных в ней веществ в клетку и выход их из нее.
Эти процессы регулируются полупроницаемостью протоплазмы, особенно ее пограничных слоев. Протоплазма проницаема для воды и в различной мере проницаема или вовсе не проницаема для растворенных в ней веществ, включая газы (подробнее об этом процессе см. на стр. 68).
Проникновение веществ в живую клетку и выход из нее зависят от свойств протопласта. При полной его проницаемости, как и при совершенной непроницаемости, клетка не могла бы жить.
Необходимо отметить избирательный характер проницаемости живой протоплазмы. Нередки случаи, когда количество вещества, поглощенного корнями из разбавленных растворов, выше, чем из растворов более концентрированных.
Иногда говорят о полупроницаемости клетки. Следует иметь в виду, что в этих случаях речь идет только о протоплазме. Клеточная оболочка обычно легко проницаема для воды и растворов солей; для коллоидных растворов с крупными частицами дисперсной фазы оболочки клеток непроницаемы, но такие частицы могут перемещаться из клетки в клетку через перфорации в ситечках ситовидных трубок (стр. 125) и по плазмодесмам (стр. 97, рис. 74).
Движение протоплазмы. Голая протоплазма и протоплазма, находящаяся внутри твердой оболочки, в живых клетках всегда находятся в движении. Движение голых протопластов происходит, как у амеб, с помощью образования псевдоподиев (выступов протоплазмы) и втягивания их или же осуществляется посредством работы жгутиков - тонких, упругих и гибких плазматических придатков тела. Классическими примерами движения голых протопластов являются движения слизевиков, или миксомицетов, на определенных стадиях их онтогенеза (рис. 10).
Движение протоплазмы внутри клетки. В своем движении протоплазма увлекает за собой находящиеся внутри ее органеллы и включения (пластиды, хондриосомы, капли жира и т. п.), по их перемещениям можно следить за движением протоплазмы. Как правило, не вся масса протоплазмы в клетке движется с одинаковой скоростью; слой, примыкающий к оболочке, движется наиболее медленно.
35
Различают несколько типов движения.
Круговое (ротационное) движение наблюдается в клетках, в которых весь протопласт сосредоточен в постенном слое, а срединная часть клеточной полости занята крупной центральной вакуолей. Характерной особенностью ротационного движения является то, что протоплазма движется в одном направлении, как бы вращаясь вокруг центра клетки. Можно ясно видеть круговое движение протоплазмы в листьях водных растений, особенно элодеи (Elodea) с ее тонкими листовыми пластинками, и валлиснерии (Vallisneria spiralis) (рис. 11).
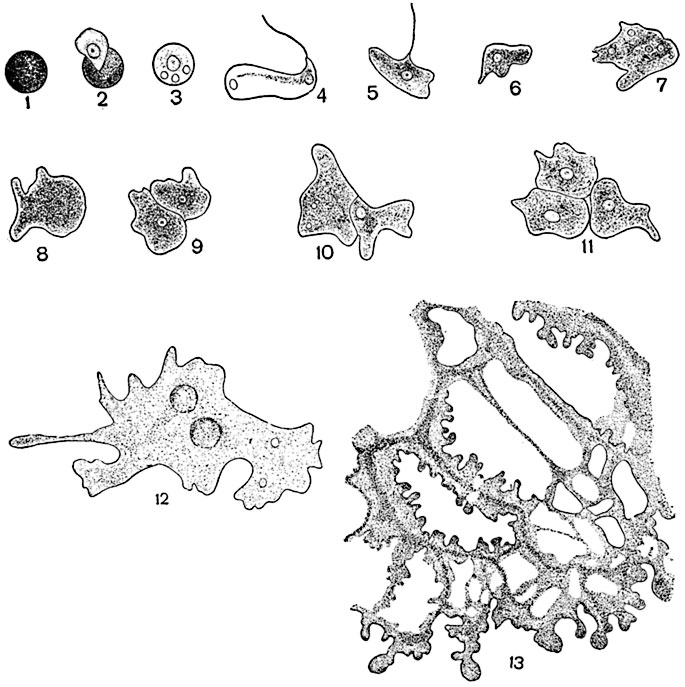
Рис. 10. Прорастание спор и образование плазмодия у слизевиков:
1 - покоящаяся спора; 2 - из ее лопнувшей оболочки выходит протопласт; 3 - протопласт, освободившийся от оболочки; 4, 5 - фазы зооспоры; 6, 7, 8 - фазы одиночной миксамебы; 9, 10 - копуляция миксамеб; 11, 12 - начальные этапы образования плазмодия; 13 - часть плазмодия слизевика Fuligo septica, ползущего по мокрой фильтровальной бумаге; почти в натуральную величину.
На продольных срезах через лист валлиснерии, особенно в теплую погоду, или на срезах, выдержанных в течение 5-10 минут в теплой воде, можно видеть, что протоплазма в ее круговом движении увлекает за собой не только хлорофилловые зерна, но порой и ядро. Круговое движение протоплазмы непостоянно по скорости и направлению и апериодично: в данной клетке оно то ускоряется, то замедляется, временами, без определенной ритмичности, приостанавливается.
Линейная скорость движения протоплазмы при ротации не высока: так, у валлиснерии при комнатной температуре она представляет величину порядка 10-20 μ в секунду, у элодеи - 10-15 μ в секунду.
36
При рассматривании в микроскоп видимая скорость движения увеличивается соответственно увеличению, даваемому микроскопом. Чрезвычайно энергично и закономерно движется протоплазма в клетках харовых водорослей (Chara и Nitella).
Струйчатое (циркуляционное) движение протоплазмы происходит в клетках, в которых протоплазма находится в виде постенного слоя и тяжей, пересекающих центральную вакуолю. Протоплазма движется многочисленными тонкими струйками по различным направлениям. Время от времени направление той или иной струйки меняется на обратное. Нередко бок о бок движутся струйки в противоположных направлениях. Тяжи в срединной области клетки медленно изменяют форму, толщину, расположение, некоторые из них постепенно исчезают, возникают новые тяжи.
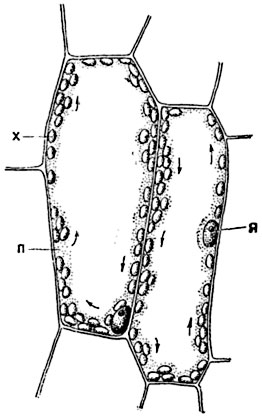
Рис. 11. Круговое движение протоплазмы в клетке мякоти листа валлиснерии (направление движения протоплазмы обозначено стрелками):
п - протоплазма; х - хлорофилловые зерна; я - ядро.
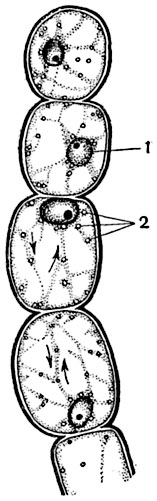
Рис. 12. Клетки волоска тычиночной нити традесканции виргинской (Tradescantia virginica); направление движения обозначено стрелками:
1 - ядро; 2 - лейкопласты.
Волоски с тычиночных нитей традесканции (Tradescantia virginica), состоящие из одного ряда клеток, представляют один из наилучших объектов для наблюдения за струйчатым движением протоплазмы (рис. 12).
Энергично происходит циркуляционное движение в волосках молодых побегов тыквы, в жгучих волосках крапивы, в молодых корневых волосках плавающих водных растений - водокраса (Hydrocharis morsus-ranae), трианеи (Trianea bogotensis).
Зависимость движения протоплазмы от внешних и внутренних условий.
Движение протоплазмы в клетках может быть первичным и вторичным. Первичное движение происходит в клетках неповрежденных и при
37
обычных для данного растения условиях. Оно имеет место у высших растений в волосках кожицы листьев и стеблей, в корневых волосках, пыльцевых трубках; из низших растений им обладают хары и другие водоросли. Вторичное движение происходит в неподвижной до того протоплазме в результате внешних воздействий - повреждения соседних клеток (как, например, при изготовлении среза), резких изменений температуры или яркости освещения. В обычных объектах лабораторной практики - листьях водных растений элодеи и валлиснерии - наблюдаются вторичные движения протоплазмы.
Резкой границы между первичными и вторичными движениями провести нельзя.
Движение протоплазмы может совершаться и в плазмолизированных клетках, хотя несколько иначе, нежели в клетках, не подвергнувшихся плазмолизу.
Движения протоплазмы стимулируются внешними условиями. Большую роль играет температура: так, для элодеи установлено, что температурные пределы для движения протоплазмы лежат между 10° С и 42° С и оптимальная (т. е. наиболее благоприятная) температура +37° С. В большинстве случаев движение протоплазмы стимулируется наличием кислорода.
Большую роль играет и освещение. Так. в листьях валлиснерии, выдержанных в темноте, освещение силой в 22,2 люкса и продолжительностью в 80 секунд возобновляет движение протоплазмы. Движение протоплазмы стимулируется либо тормозится различными веществами в окружающей среде: так относительно элодеи замечено, что движение протоплазмы в клетках значительно более энергично в листьях, взятых с экземпляра из сосуда с водой после того, как в воду добавлено несколько капель спирта.
Движение протоплазмы становится более энергичным перед началом или в течение усиленной ее деятельности (например, перед образованием придаточных побегов в клетках кожицы листа бегонии и т. д.).
38
1 От греческих "протос" - первый и "пластос" - вылепленный.
2 Термин "цитоплазма" удобен для применения в случаях, если надо подчеркнуть, что речь идет именно о протоплазме клетки (цитоплазме), а не о всем живом теле ее (т. е. не о протопласте), "протоплазме" в широком смысле слова.
3 От греческих "эктос" - внешний; "гиалос" - стекло; "лемма" - скорлупа, кожица; "эндо" - внутренний; "мезос" - средний; "тонос" - напряжение.
4 Цитологи и биохимики мембранами называют группы из правильно ориентированных молекул, образующих разграничивающие слои, отделяющие цитоплазму от ретинулюма, вакуолей и т. п.
5 Полисахарид.
6 Ф. Энгельс, Диалектика природы, Госполитиздат, 1964, стр. 264-265.
7 От греческих "колла" - клей и "идос" - вид.
8 От латинского coacervatio - собирание в кучу, накопление.
9 Эластичностью тела называется способность после деформации принимать исходную форму.
10 Микроманипулятор - прибор, при помощи которого можно изолировать от организма группу клеток, клетку и даже часть ее содержимого. Микроманипулятор обычно бывает соединен с микроскопом.