

СТРОЕНИЕ МНОГОЛЕТНИХ СТЕБЛЕЙ ДРЕВЕСНЫХ РАСТЕНИЙ
У растений, наземные части которых живут в течение многих лет, камбий функционирует периодически. Многолетняя деятельность камбия приводит к коренным изменениям в строении древесины и луба. Эти изменения обычно называют вторичными.
Вторичная ксилема, или вторичная древесина
Строение древесины хвойных. В качестве примера возьмем древесину сосны обыкновенной (Pinus silvestris). Она состоит почти сплошь из трахеид с окаймленными порами (рис. 135). Трахеиды расположены радиальными рядами; кое-где радиальные ряды удваиваются, в соответствии с увеличением числа клеток камбиального кольца. Трахеиды в большинстве имеют на поперечном разрезе почти прямоугольные очертания. В древесине, образовавшейся за один год деятельности камбия, т. е. в пределах ее годичного кольца, трахеиды различны: одни из них - ранние трахеиды, образовавшиеся в результате весенней и раннелетней работы камбия, - имеют в радиальном направлении размер больший, нежели в тангентальном. Стенки этих трахеид слабо утолщены, и просвет сравнительно велик. Радиальные стенки их снабжены порами, расположенными в один продольный ряд; окаймление пор резко выражено; в замыкающей пленке вокруг торуса имеются мельчайшие перфорации1. Концы ранних трахеид закруглены. В трахеидах поздней древесины, образованной камбием в конце лета и осенью, радиальный размер значительно меньше тангентального; оболочка сильно утолщена, а клеточный просвет мал. Поры узки, малочисленны, с окаймлениями, почти не выступающими над внутренней
155
поверхностью клеточной стенки. В самых поздних трахеидах поры иногда находятся не в радиальных, а в тангентальных стенках.
Трахеиды ранней древесины в соответствии с их строением являются преимущественно элементами проводящей системы; поздние же трахеиды, по строению принадлежащие к типу волокнистых трахеид, функционируют в большей мере как механические элементы.
Среди продольных трахеид проходят древесинные лучи и смоляные ходы - вертикальные и горизонтальные.
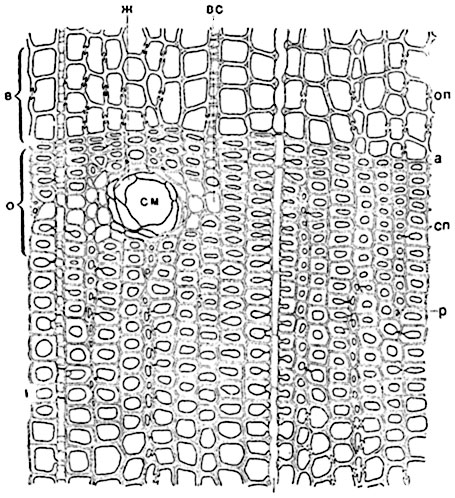
Рис. 135. Поперечный разрез древесины сосны (Pinus silvestris):
а - граница между двумя годичными слоями; о - осенняя древесина; в - весенняя древесина; оп - окаймленная пора трахеиды; сп - срединная пластинка в оболочке трахеид; р - место удвоения к радиальном ряде трахеид; см - смоляной ход; ж - эпителий смоляного хода; вс - молодой древесинный луч.
Древесинные лучи у сосны двоякого рода: а) узкие и б) широкие, со смоляными ходами, в средних ярусах состоящие из нескольких рядов клеток. Верхние и нижние ярусы широкого луча однорядны.
Древесинные лучи идут радиально в направлении от древесины к камбию и продолжаются по другую сторону камбия в лубяные лучи. Древесинные лучи сосны строятся из клеток двух типов, что ясно видно на продольных разрезах ствола (рис. 136, 137). Средние ярусы клеток луча состоят из живых паренхимных клеток, богатых крахмалом; оболочки этих клеток слабо одревесневают; вертикальные и горизонтальные их стенки снабжены крупными простыми порами. Верхний и нижний ярусы луча заняты клетками с одревесневшими оболочками, имеющими во всех стенках мелкие окаймленные поры. Это лучевые трахеиды древесинного луча; после сформирования они теряют живое содержимое; их главная функция - проведение воды с минеральным раствором в направлении радиуса ствола.
Клетки древесинных лучей сообщаются с примыкающими к ним продольными трахеидами посредством пор, окаймленных со стороны трахеид; со
156
стороны луча поры в лучевых трахеидах мелкие, окаймленные, а в клетках средних ярусов, содержащих крахмал, крупные, угловатые в очертании, слабо и частично окаймленные. В широких лучах срединная, расширенная часть содержит горизонтальный смоляной ход.
Смоляные ходы - длинные каналы, заполняющиеся смолой. У сосны они выстланы тонкостенными паренхимными клетками - эпителием,
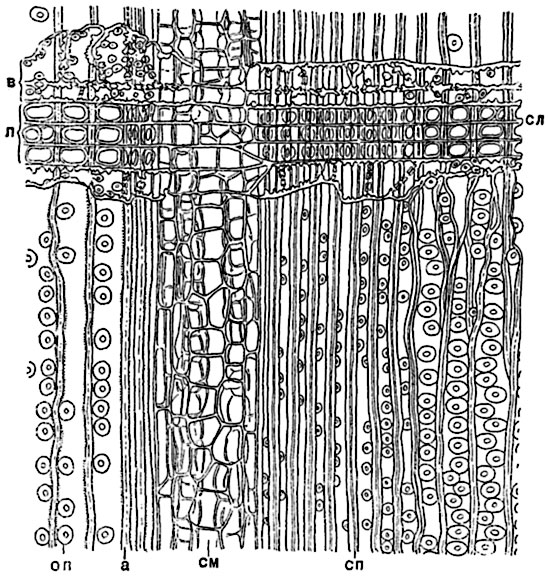
Рис. 136. Продольный радиальный разрез древесины сосны (Pinus silvestris):
а - граница между двумя годичными слоями; оп - окаймленные поры на радиальных стенках трахеид, в плане; сп - срединная пластинка в оболочках трахеид; см - смоляной ход; сл - древесинный луч, в нем: в - лучевые трахеиды, п - клетки с пластическими веществами.
который представляет собой особую ткань, выделяющую смолу. Как вертикальные, так и горизонтальные смоляные ходы сосны имеют схизо-лизигенное происхождение.
Смоляные ходы луча сообщаются с вертикальными смоляными ходами, благодаря чему все смоляные ходы образуют единую систему.
В древесинных лучах имеются узкие радиальные воздухоносные межклетники, продолжающиеся в лубяных лучах. На 1 мм2 тангентального сечения древесины сосны число древесинных лучей (частота лучей) превышает 30. На благоприятной для сосны почве и при хорошем освещении частота лучей повышается.
У всех хвойных трахей нет, а трахеиды иных типов, кроме пористых,- кольчатые и спиральные, имеются только в первичной древесине. У некоторых хвойных (у тисовых, Pseudotsuga) трахеиды древесины имеют дополнительные утолщения стенок в
157
виде спиральных лент. Перфорации в замыкающих пленках пор у лиственницы и у секвойи столь крупны, что видимы в микроскоп. Продольная паренхима не только у обыкновенной сосны, но и у всех видов рода Pinus представлена в древесине только клетками эпителия смоляных ходов; у многих хвойных (в том числе у елей, лиственниц) продольная древесинная паренхима образуется независимо от смоляных ходов ежегодно в конце деятельности камбия; количество ее, однако, весьма мало. У некоторых хвойных имеются только узкие древесинные лучи. У многих (в том числе у пихт, Pseudolarix) лучи состоят нормально из однотипных (паренхимных) клеток, а после поранения в их лучах появляются трахеидальные клетки. Смоляные ходы в здоровой древесине встречаются только у четырех родов голосеменных: сосен, елей, лиственниц и псевдотсуги. У остальных родов в здоровой древесине смоляные ходы отсутствуют.
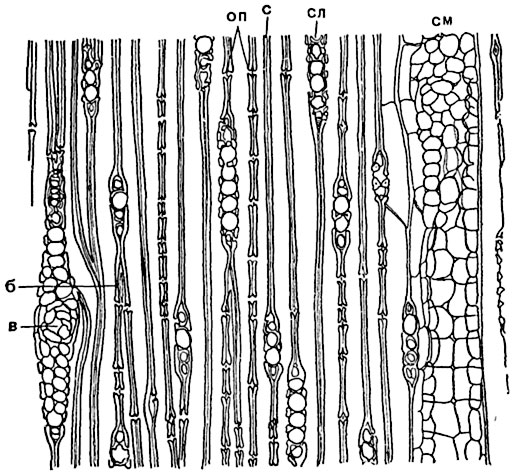
Рис. 137. Продольный тангентальный разрез древесины сосны:
в - конец трахеиды; оп - окаймленные поры в разрезе; с - срединная пластинка в общей стенке двух трахеид; см - продольный смоляной ход; б - смоляной ход в древесинном луче; сл - древесинный луч.
Строение древесины двудольных. Древесина двудольных по своему составу гораздо разнообразнее древесины голосеменных. В ней можно различить проводящие, запасающие, механические и выделительные элементы.
В древесине двудольных проводящие элементы представлены трахеями и трахеидами, чаще всего пористыми, точечными, реже сетчатыми и лестничными. У некоторых растений (в том числе у кленов, лип, виноградной лозы) все точечные трахеи и трахеиды или часть их имеют дополнительные внутренние утолщения стенок в виде спиралей, реже в виде колец или сеточки (рис. 138). В перегородках между члениками трахей имеется простая или множественная перфорация (рис. 69).
При множественной перфорации перфорационные пластинки могут быть: а) лестничными - с несколькими или многими узкими параллельными перфорациями (рис. 103) или б) сетчатыми - с системой перекладин между перфорациями, имеющей вид сетки. Диаметр трахей представляет величину от нескольких сотых до нескольких десятых долей миллиметра: у березы - до 0,085 мм, у дуба - 0,2-0,3 мм, у некоторых лиан - до 0,5-0,6 мм. Длина трахеи с трудом поддается измерению.
158
Проводящая система древесины обслуживает в основном восходящий ток, транспортируя в направлении от корней к стеблю с его ответвлениями и листьями воду с раствором минеральных веществ, всасываемую корнями; по этой же системе весной до распускания листьев подается в набухающие почки раствор пластических веществ (главным образом сахаров), образующихся при мобилизации зимних запасов, накопленных в тканях запасающей системы.
У большинства двудольных в древесине имеются и трахеи и трахеиды; очень немногие (в том числе Drimys, из семейства магнолиевых) имеют
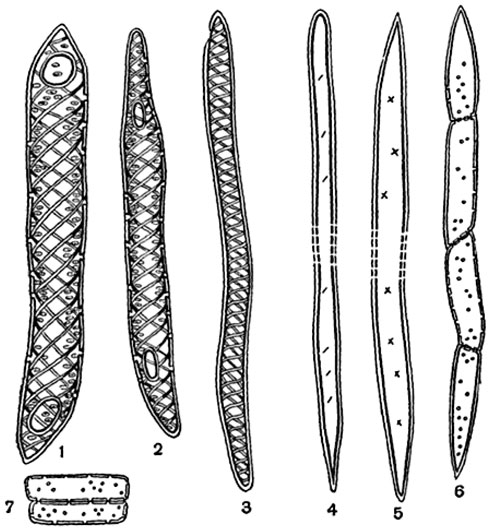
Рис. 138. Элементы древесины липы (Tilia cordata) (мацерированный материал):
1, 2 - членики спирально-пористых сосудов; 3 - трахеида со спиральным утолщением; 4, 5 - либриформ; 6 - тяж древесинной паренхимы; 7 - клетки сердцевинных лучей.
только трахеиды, некоторые (ясени, платаны) - только трахеи. В размещении сосудов на поперечном сечении годичного кольца древесины различают два основных варианта: а) одна зона кольца прироста сильно отличается от другой по диаметру, или по численности сосудов, или по обоим этим показателям; такую древесину называют кольцесосудистой; б) в пределах годичного кольца диаметр и частота сосудов не обнаруживают резких отличий, такая древесина называется рассеяннососудистой. Образцом кольцесосудистой древесины может служить ясень, рассеяннососудистой - ольха (рис. 139).
Запасающие элементы в древесине двудольных представлены древесинной паренхимой. Клетки продольной древесинной паренхимы располагаются цепочками. Каждая цепочка, или тяж древесинной паренхимы, образуется из прозенхимной клетки молодого прироста путем деления
159
ее поперечными перегородками. Из клеток тяжа две крайние сохраняют заострение на конце (рис. 138). Взрослые клетки древесинной паренхимы имеют обычно умеренно утолщенную одревесневшую оболочку с простыми круглыми порами и живое содержимое в виде постенного слоя протоплазмы с ядром и лейкопластами. В клетках накопляется на зиму масло (у лип, берез) или крахмал (у буков). Весной запасы обычно мобилизуются и перемещаются к распускающимся почкам и к камбию2.
Поры в оболочках паренхимных клеток особенно крупны в стенках, примыкающих к сосудам, и мелки и малочисленны или вовсе отсутствуют в стенках, граничащих с древесинными волокнами.
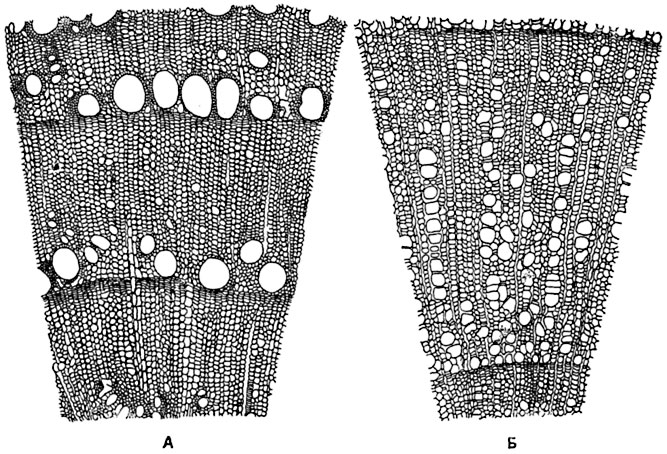
Рис. 139. Частичные поперечные разрезы древесины:
А - кольцесосудистой (ясеня Fraxinus excelsior); Б - рассеяннососудистой (ольхи клейкой Alnus glutinosa).
Если при дифференциации прозенхимной клетки из камбия она остается живой, оболочка ее получает простые круглые поры и разделения клетки поперечными перегородками не происходит, то образуется веретеновидная клетка древесинной паренхимы3.
Механические элементы древесины представлены склеренхимой; склеренхиму древесины называют либриформом, а ее клетки - древесинными волокнами (рис. 138). Древесинные волокна обладают чертами строения, характерными для всякой склеренхимы. Надо только добавить, что клетки либриформа относятся к числу наиболее коротких растительных волокон4 и что стенки их всегда одревесневают. Древесинными волокнами, не обладающими всеми чертами типичной
160
склеренхимы, являются живые древесинные волокна без перегородок и перегородчатые. У живых древесинных волокон стенки слабо утолщаются, а в клеточной полости в течение ряда лет сохраняется живое содержимое и периодически накопляются и мобилизуются запасные вещества (крахмал, масло)5. В волокнах перегородчатого либриформа по завершении их роста в длину и утолщения стенок образуются
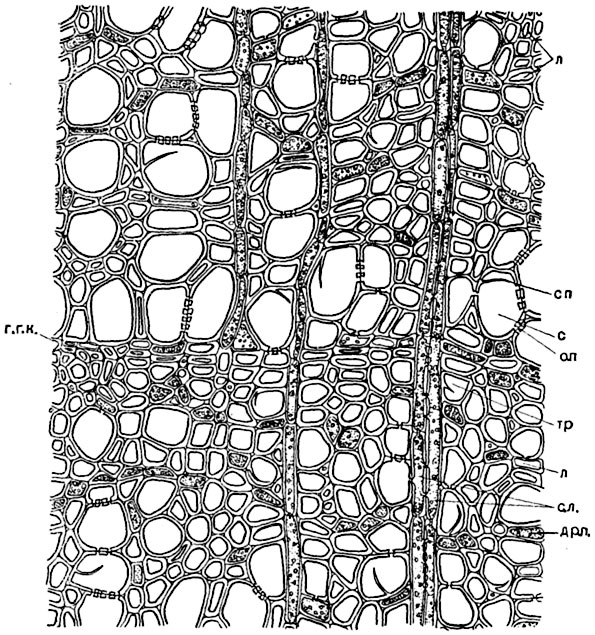
Рис. 140. Поперечный срез древесины липы:
г. г. к. - граница годичного кольца; с - спирально-пористый сосуд; сп - обрывок спирального утолщения оболочки; о. п. - окаймленная пора на стенке сосуда; тр - трахеида с обрывком спирального утолщения; др. п. - древесинная паренхима; л - либриформ; с. л. - сердцевинный луч.
поперечные перегородки - весьма тонкие и неодревесневающие. Такие волокна имеются, например, у виноградной лозы, плюща.
Выделительные элементы в древесине представлены обычно клетками с отложениями кристаллов щавелевокислого кальция. У очень немногих видов (в том числе у дынного дерева Carica papaya и других представителей тропического семейства Papayaceae) в древесине образуется система млечников.
Древесинные лучи двудольных (рис. 140, 141, 142) весьма разнообразны по конфигурации и по размерам. У некоторых видов
161
имеются только узкие лучи, у иных (у дубов) - и узкие и широкие6. Нередки агрегатные лучи (у ольхи, лещины, граба)7. Лучи могут быть гомогенными (однородными) и гетерогенными (разнородными). В гетерогенных лучах (у ив) имеются лежачие и стоячие клетки.
Воздухоносные межклетники образуются обычно между лежачими клетками. С сосудами и трахеидами, примыкающими к лучу, сообщаются
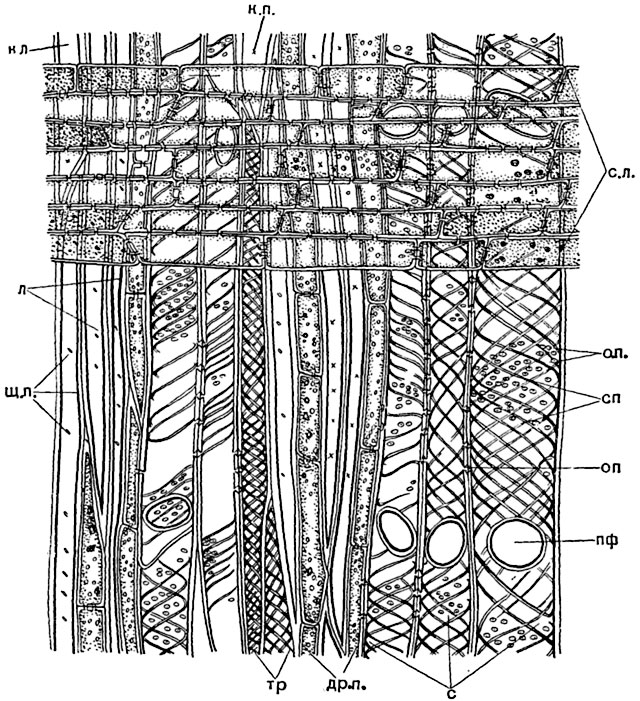
Рис. 141. Продольный радиальный срез древесины липы:
с - спирально-пористые сосуды; пф - перфорация сосуда; сп - спиральные утолщения стенок сосудов; о. п. - окаймленные поры; тр - трахеиды со спиральными утолщениями; л - либриформ; к. п. - крестовидная пора; щ. п. - щелевидная пора; др. п. - древесинная паренхима; с. л. - сердцевинный луч.
посредством пор только стоячие клетки. Лежачие клетки сообщаются порами между собой и со стоячими клетками. Ряды лежачих клеток обслуживают передвижение веществ в радиальном направлении, а стоячие обеспечивают сообщение лежачих клеток с сосудами.
Сезонные различия в древесине. У растений умеренных и холодных
162
широт древесина, порождаемая камбием в течение одного вегетационного периода, неединообразна гистологически: ранняя древесина, образующаяся весной и в начале лета, относительно богата широкопросветными сосудами, а в древесине поздней, появляющейся в конце лета и осенью, более сильно представлены элементы механической и запасающей систем, сосудов в поздней древесине мало, и они сравнительно узкопросветны. Особенно велико различие между ранней и поздней древесиной у кольцесосудистых пород (рис. 139).
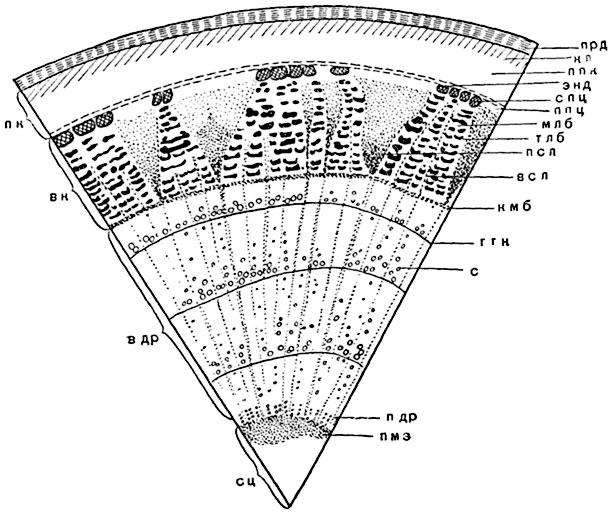
Рис. 142. Схема строения многолетней ветки липы (Tilia cordata):
прд - перидерма; кл - колленхима; ппк - паренхима первичной коры; энд - эндодерма; спц - склеренхима перицикла; ппц - паренхима перицикла; млб - мягкий луб; тлб - твердый луб; псл - первичный сердцевинный луч; всл - вторичный сердцевинный луч; кмб - камбий; ггк - граница годичного кольца древесины; с - сосуды; пдр - первичная древесина; вдр - вторичная древесина; пк - первичная кора; вк - вторичная кора; сц - сердцевина; пмз - перимедуллярная зона.
В связи с сезонными различиями в приросте древесины стоит наличие зон прироста, различаемых на поперечных разрезах древесины в виде концентрических колец (рис. 135, 139, 142); на продольных радиальных разрезах границы между зонами (кольцами) видны как почти прямые параллельные линии. Обычно каждое кольцо образуется в один вегетационный период, и кольца прироста могут быть названы годичными кольцами.
Границы между годичными приростами древесины видны для нас в силу того, что гистологические различия между ранней и поздней древесиной вызывают и различия оптические - в цвете и блеске. В пределах одного годичного слоя переход от древесины ранней к поздней более или менее постепенен. На грани же между двумя годичными слоями, где к наиболее типично выраженной поздней древесине одного года непосредственно примыкает ранняя древесина последующего года, различие резко выражено и
163
обычно настолько бросается в глаза, что на поперечных разрезах древесины мы видим непосредственно или, реже, с помощью лупы как бы вычерченную пограничную линию - круг или овал.
Число колец прироста древесины при основании ствола дерева указывает на возраст деревьев умеренных и высоких широт. Это правило имеет исключения: при некоторых обстоятельствах происходит удвоение, при других выпадение кольца. Удвоение кольца прироста древесины происходит при двукратном в один вегетационный период зазеленении кроны дерева: отмечалось, например, у рябины, дуба летнего и липы мелколистной образование двух различимых глазом колец прироста в году, когда деревья, будучи объедены в июне гусеницами, вновь покрылись в том же году листвой8. Удвоение кольца может происходить и в том случае, если молодая листва отмирает при весенних заморозках, а после того распускаются спящие почки. Кольца, появляющиеся в результате удвоения (ложные кольца), имеют менее резко выраженные границы, чем настоящие годичные кольца, и, кроме того, граница обычно не образует полного круга.
Выпадение кольца прироста происходит в наиболее слабых побегах угнетенных деревьев в естественных условиях и в культуре. Выпадает целая серия годичных колец прироста при основании ветвей некоторых "плакучих" разновидностей древесных пород, при основании свисающих ветвей ели. У карликовых деревьев, выращиваемых в горшках со скудной и слабо увлажняемой почвой, выпадают многие кольца прироста.
Дифференцировка древесины по зонам одного слоя прироста (годичного или негодичного) является отчасти результатом воздействия среды на данное дерево, отчасти наследственным свойством, осуществляющимся при различных условиях воздействия внешней среды. Обнаружено, что далеко не у всех пород существует соответствие между периодичностью или апериодичностью метеорологических условий в течение года и характером образующейся древесины. У некоторых пород такое соответствие имеет место; так, например, у особей тикового дерева (Tectona grandis), растущих в довольно постоянном круглый год климате в Бейтензорге, на Западной Яве, древесина гомогенна и колец прироста не видно, а у особей того же вида, живущих в условиях климата, изменяющегося по сезонам (как в районе Восточной Явы, где чередуются сухое и дождливое времена года), древесина не гомогенна и в ней ясно заметны кольца прироста. С другой стороны, некоторые тропические виды (в том числе туна Toona serrata, T. sureni) даже в условиях ровного климата Бейтензорга образуют древесину с ясно выраженными кольцами прироста, а иные (Plumiera acuminata), живя в условиях климата Восточной Явы, где в сухое время года они даже теряют листву, не обнаруживают в древесине дифференцировки на зоны широкопросветных и узкопросветных тканей, и колец в древесине не заметно.
Уже давно замечено, что деятельность камбия находится в тесной связи с развитием почек. Изучение хода весеннего пробуждения камбия с помощью анатомического исследования продольно-тангентальных полосок, отпрепарированных с поверхности древесинного тела, показало точно, что возобновление деятельности камбия после зимнего покоя начинается под набухающими почками и распространяется отсюда вниз и почти одновременно с этим по окружности ствола. Весеннее "пробуждение" камбия у пород с кольцесосудистой древесиной (у ясеней, дубов, вязов, каштана) продвигается очень быстро и достигает основания ствола в несколько дней. У видов с рассеяннососудистой древесиной дело идет значительно медленнее: у
164
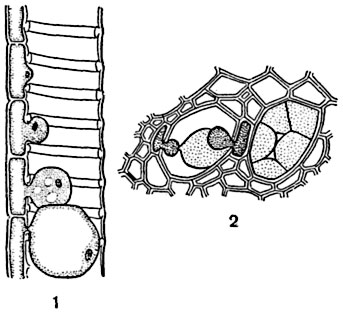
Рис. 143. Сосуд с тиллами (белая акация Robinia pseudacacia):
1 - продольный срез; 2 - поперечный срез.
березы, бука камбий функционирует в течение нескольких недель только у основания почек; лишь после того как развернутся листья, начинается образование новой древесины в молодых веточках, а затем уже в ветвях и в стволе. У хвойных "пробуждение" камбия распространяется по ветвям и стволу очень быстро, особенно в условиях хорошего освещения, однако в нижних ветвях, а у сильно затененных деревьев и в стволе процесс возобновления камбиальной деятельности продвигается медленнее.
Вопрос о характере зависимости между распусканием почек и возобновлением деятельности камбия был выяснен физиологическими исследованиями. Многочисленные эксперименты показали, что деятельность камбия обусловливается наличием почек и передвижением из них некоторых веществ в камбий.
Последующие изменения во взрослой древесине. Трахеи, реже трахеиды, закупориваются со временем тиллами - отрогами клеток древесинной паренхимы, внедряющимися в полость сосудов через тонкие участки их стенок (т. е. через поры, рис. 143). Паренхимная клетка образует одну или несколько тилл; тиллы продолжают расти и в полости сосуда, формируясь в тонкостенные пузыри или мешки, содержащие постенный слой протоплазмы, иногда с крахмальными зернами. В протоплазме тиллы иногда имеется и клеточное ядро, перешедшее в нее после деления ядра паренхимной клетки, произведшей тиллу; в этих случаях тилла может обособляться и образовывать делением новые тиллы в полости сосуда. Входя в соприкосновение друг с другом, тиллы принимают многогранную форму и нередко, срастаясь оболочками, образуют внутри сосуда ложную паренхимную ткань. Со временем тиллы отмирают; к этому моменту у них в некоторых случаях сильно утолщаются и одревесневают оболочки. В нормальных случаях тиллы образуются или уже на первом-втором году существования сосуда (во многих сосудах древесины белой акации), или позже9. У некоторых двудольных (у вишен, берез, кленов) тиллообразования не происходит: сосуды перестают функционировать в силу заполнения их просветов минеральными или органическими веществами.
Тиллы в древесине как поделочном материале представляют нередко отрицательное явление: они препятствуют пропитыванию древесины антисептиками; это особенно дает о себе знать у бука, в древесине которого уже после рубки дерева могут быстро закупориваться тиллами сосуды, ранее бывшие открытыми.
Трахеиды хвойных перестают проводить воду с раствором в силу того, что замыкающие пленки их окаймленных пор занимают положение, при котором торус закрывает отверстие канала поры. Полости трахеид иногда заполняются живицей - раствором смол в эфирных маслах.
Живые паренхимные клетки древесины рано или поздно отмирают и перестают служить хранилищем запасов. В первую очередь отмирают клетки
165
продольной паренхимы и живые волокна; поперечная паренхима в древесинных лучах живет дольше. Перед отмиранием клеток усиливается одревеснение их оболочек и снижается содержание воды в оболочках и полостях клеток. В гистологических элементах всех родов может происходить со временем отложение консервирующих веществ, нередко с интенсивной окраской. Более старая часть древесины ствола у многих пород резко выделяется цветом; ее называют ядром в отличие от окружающей ее более молодой части древесины - заболони.
Из особенно характерных окрасок ядра отметим: желтую у барбариса и у бересклета европейского, красную у кипарисов, темно-фиолетовую или почти синюю у кампешевого дерева (Haematoxylon campeschianum из семейства бобовых), черную у эбеновых деревьев (индийских и западноафриканских видов рода Diospyros). У многих древесных пород (в том числе у буков, груш, кленов) гистологические элементы ядра заполняются углекислым кальцием. В полостях клеток ядра (и отчасти спелой древесины10) откладываются органические вещества - эфирные масла, смолы, камеди, дубильные вещества и иные. Коричневый цвет ядра дубов обусловливается, например, бурым веществом, выпадающим в твердом виде после окисления дубильных веществ.
Ядровые вещества появляются обычно в живых клетках сердцевины и древесинной паренхимы, а из них проникают в другие элементы ядровой древесины; эти вещества заполняют полости клеток, а иногда и пропитывают клеточные оболочки, адсорбируясь ими.
В ядровой древесине кевового дерева (Pistacia mutica) и белой акации паренхимные клетки могут долго сохраняться живыми, причем содержимое их (крахмал, щавелевокислый кальций) подвергается сезонным изменениям.
У некоторых древесных пород (у ив, тополей) более старая древесина становится мягкой и легкой и часто разрушается грибами, проникающими в нее через трещины, образующиеся, например, при обломе ветвей; в стволе образуется дупло.
Древесина находит самое разнообразное применение. Она служит, во-первых, топливом. Наиболее калорийны древесины с большим относительным количеством клеток с толстыми, сильно одревесневшими стенками (у дуба, бука, сосны). В большом количестве древесина идет на постройки, мебель, шпалы, телеграфные столбы, аппаратуру и т. д. В каждом случае сообразуются со строением древесины и с ее физико-механическими свойствами. Например, для частей машин, мельниц ценится прочная и твердая древесина граба, бука. Для изготовления коробов, для драни подходит легко раскалываемая древесина елей, осин. Древесины дубов, белой акации, сосуды которых рано и полно закупориваются тиллами, пригодны для бочек под пиво и спирт. Древесины, бедные лигнином, смолами и камедями, минеральными веществами и пигментами, притом обладающие длинными и мягкими волокнистыми элементами (осина, липа, ель), дают хороший материал для получения целлюлозной массы, служащей для производства бумаги, искусственных тканей, для осахаривания в глюкозу и затем для получения спирта и, наконец, синтетического каучука.
Твердая, плотная, прочная и красиво окрашенная ядровая древесина (дуба, черного дерева и т. д.) представляет ценный материал для красивой мебели.
При сухой перегонке древесины получаются разнообразные ценные продукты: метан, уксусная кислота, древесный спирт, ацетон, деготь (смола).
166
Из дегтя извлекаются разнообразные вещества: из букового дегтя, например, получают важные дезинфицирующие средства - креозот, гваякол.
167
1 Наличие их обнаруживается в опытах фильтрации воды с тушью. Под микроскопом эти перфорации не видны.
2 У пород, начинающих плодоносить поздно и в дальнейшем плодоносящих не каждый год, в голы без плодоношения мобилизуется лишь небольшая доля запасов. Это относится, например, к букам, зацветающим в возрасте 40-50 лет и приносящим обильно плоды в среднем раз в 5 лет.
3 Раньше такие клетки называли "заменяющими волокнами" или "замещающими волокнами".
4 Длина древесинных волокон колеблется в пределах 0,3-1,7 мм.
5 У некоторых древесных пород (у вязов, шелковицы, орешников и других) внутренний слой оболочки живых древесинных волокон образуется из гемицеллюлоз, служащих запасными веществами: к зиме отлагающихся, а весной гидролизующихся.
6 У дубов широкие древесинные лучи достигают в высоту 5-8,5 мм.
7 Агрегатным (иначе ложношироким или ложным) лучом называют группу узких древесинных лучей, представляющуюся глазу или при малом увеличении в виде одного широкого луча.
8 В экспериментах с удалением распустившейся весной листвы рябины, орешника и конского каштана, после чего деревца вновь покрылись листвой, искусственно вызывалось появление за год двух колец прироста древесины; на границе между ними образовалась прослойка клеток, чрезвычайно тонкостенных.
9 Быстрое закупоривание сосудов тиллами происходит при поранениях, происходящих при опадении листьев или веток, при обламывании или обрезании ветвей, при черенковании.
10 В дендрологии и лесотехнике спелой древесиной принято обозначать ту внутреннюю часть ствола, которая не отличается по окраске от заболони, но содержит (после срубки) меньшее количество воды и выделяется при спиливании в виде более сухого участка.