

СОСТАВ И СТРОЕНИЕ ОБОЛОЧКИ
Целлюлоза. Оболочка молодых клеток у всех растений, за исключением грибов1, обычно состоит в основном из целлюлозы (клетчатки) - углевода эмпирической формулы (C6H10O5) n. Оболочка состоит из веществ в состоянии геля; она нерастворима в воде, но способна впитывать воду и растворенные в ней вещества и при этом несколько набухать. Оболочка анизотропна, т. е. показатели ее физических свойств (светопреломления, упругости и т. д.) неодинаковы по различным направлениям. Детали строения сложной молекулы целлюлозы, а следовательно, и оболочки выяснены на основе целого комплекса исследований биохимических и оптических - с помощью оптического, поляризационного и электронного микроскопов и лучей Рентгена.
Целлюлоза состоит из многих длинных, напоминающих цепочки молекул, образуемых повторяющимися единицами, состоящими из двух глюкозных остатков (целлобиоза), повернутых один относительно другого на 180° и соединенных глюкозидными связями между первым и четвертым атомами (рис. 52). Эта структурная формула целлюлозы общепринята. Однако ее строение определяется взаимным расположением длинных молекул (макромолекул).
Первое представление о том, что целлюлозные волокна построены из длинных образований, было дано Негели.
Изучая морфологическое строение целлюлозных волокон растений, он пришел к заключению, что волокна образованы из удлиненных субмикроскопических единиц - кристалликов, которые назвал мицеллами.
Согласно современной мицеллярной теории, целлюлозное волокно - макромолекула - состоит из упорядоченных областей мицелл, где цепи ориентированы параллельно оси волокна и прочно удерживаются межмолекулярными силами, и их неориентированных областей - аморфной части, где довольно легко может происходить взаимодействие целлюлозы с другими веществами. Длинные цепи проходят как через ориентированные участки, так и через неориентированные. Одна молекулярная цепь может участвовать в образовании нескольких мицелл. Длинные цепи прерываются внутри мицелл. На рисунках 53, 54 изображены схемы мицеллярного строения целлюлозы, которые иллюстрируют новые мицеллярные представления.
82
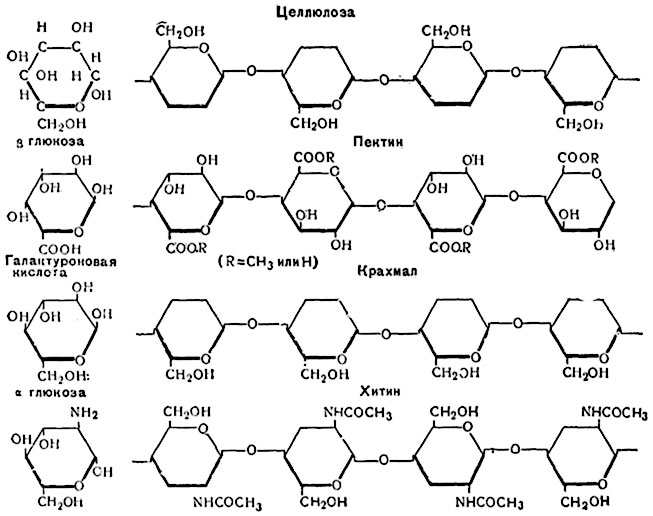
Рис. 52. Строение молекул целлюлозы, пектина, крахмала, хитина.
Рентгенографическим методом определено, что средняя длина мицеллы около 600 А°, тогда как длина одного остатка глюкозы 5,1-5,2 А°, а целлобиозы 10,3 А°, а средняя ширина мицеллы находится в пределах 50-100 А° (ширина целлобиозы - 7,9-8,35 А°).
Макромолекула целлюлозы содержит 30 000 или более групп целлобиозы. Молекулярный вес природной целлюлозы считают не меньшим 570 000.
Многие исследователи по-разному располагают элементарные единицы в кристаллической ячейке природной целлюлозы. Это, по-видимому, зависит как от различий в методах исследований, так и от того, что безводная целлюлоза существует в нескольких модификациях.
Макромолекулы целлюлозы собраны в пучки, которые часто называют фибриллами. Химическая структура целлюлозы, по-видимому, определяет фиброидную (как волокнистую, так и сетчатую) структуру толщи клеточной оболочки.
Каждая микрофибрилла имеет диаметр 1/50-1/30 μ.
Физические и химические свойства клетчатки объясняют, почему волоски и волокна, стенки которых состоят из клетчатки, употребляют для приготовления ткани. Клетчатка характеризуется большой прочностью, зависящей от упругости и эластичности. Волокна в полотняной ткани можно перегибать сколько угодно раз, и они не ломаются; таких изгибов не выдержит стальная пластинка. Одновременно с этим клетчатка проявляет и большую химическую стойкость. В отличие от крахмала она не изменяется от действия горячей воды, в которой она не набухает в такой мере, как крахмал.
Целлюлоза относится к индифферентным веществам: она не является ни кислотой, ни щелочью. Она стойка к температурным воздействиям и
83
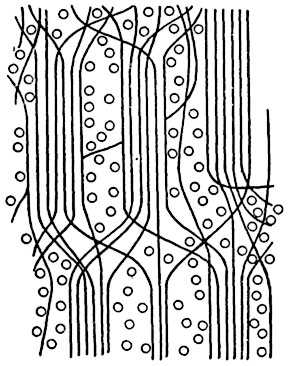
Рис. 53. Схема мицеллярного строения целлюлозы.
Длинные молекулы (цепочки) целлюлозы соединяются в пучки - мицеллы. Длинные молекулы клетчатки (около 1,5 μ), обозначенные черными линиями, могут проходить через несколько мицелл. Молекулы воды, насыщающие оболочку, обозначены кружками.

Рис. 54. Схема мицеллярного строения целлюлозы в лубяном волокне в поперечном и продольном сечениях (по Фрей-Висслингу).
Мицеллы (белые) и междумицеллярные пространства (черные) образуют стройную систему; пунктирные прямоугольники - участки правильно ориентированных мицелл.
может быть нагрета без разложения до 200° С и несколько выше, нерастворима в обычных химических растворителях и противостоит и щелочам, и слабым кислотам даже при кипячении.
Без разрушения мицелл целлюлоза растворяется только в реактиве Швейцера - растворе гидрата окиси меди в крепком аммиаке. После обработки серной кислотой целлюлоза переходит в особое амилоидное видоизменение, окрашиваемое йодом в синий цвет. Подобное же окрашивание ее получается при действии хлор-цинк-йода.
Клетчатка (целлюлоза) имеет большое значение в промышленности и технике. Обрабатывая клетчатку щелочью и сероуглеродом, получают искусственный шелк, называемый вискозой. После растворения клетчатки в аммиачном растворе гидрата окиси меди и дополнительной обработки получают так называемый медноаммиачный шелк, представляющий чистую целлюлозу. Искусственный шелк получают еще, обрабатывая клетчатку смесью азотной и серной кислот, в результате чего получается нитроклетчатка; последнюю растворяют в смеси спирта с этиловым эфиром и подвергают обработке. Ацетилклетчатку получают путем обработки клетчатки уксусной кислотой; после дополнительной обработки получают ацетатный шелк. Один из приемов получения искусственного шелка состоит в продавливании густого раствора клетчатки через тонкое сито в подкисленную воду. Из ацетилклетчатки приготовляют пластические материалы, применяемые, например, для изготовления кинолент.
Путем обработки клетчатки азотной кислотой получают нитроклетчатку, а после сплавления последней с камфарой - целлулоид. Нитроклетчатка, как и ацетилклетчатка, может быть использована для изготовления лаков высокого качества. Применение нитроцеллюлоз в технике и оборонном деле общеизвестно.
Тринитроклетчатка (пироксилин) используется в качестве взрывчатого вещества, особенно в горном деле.
Клетчатка используется для получения глюкозы путем гидролиза. Осахаривание целлюлозы производится, например, действием 0,5%-ного раствора H2SO4 при 160° С под высоким давлением (в автоклаве). Глюкоза может служить исходным материалом для получения этилового спирта. Этиловый спирт используется как сырье для получения синтетического каучука по методу С. В. Лебедева.
Источником чистой клетчатки может быть и древесина.
Целлюлоза разрушается действием определенных ферментов, содержащихся в
84
теле некоторых организмов из числа простейших животных (Protozoa), бактерий и грибов.
В пищеварительном аппарате травоядных животных (лошадей, коров и др.) ферменты, подготовляющие клетчатку к усвоению, выделяются микроорганизмами (бактериями, простейшими).
В состав клеточной оболочки нередко входят вещества, относящиеся к гемицеллюлозам (полуклетчаткам); это твердые полисахариды двух видов: гексозаны (C6H10O5) n и пентозаны (C5H8O4) n. При гидролизе гексозаны (галактаны, маннаны) дают сахара гексозы (C6H12O6), пентозаны же (арабан, ксилан) - сахара пентозы (C5H10O5).
Гемицеллюлозы, подобно клетчатке, имеют очень длинные молекулы. Гемицеллюлозы по отношению к химическим агентам значительно менее стойки, нежели целлюлозы: они гидролизуются с образованием Сахаров в слабых растворах кислот (например, в кипящей 1%-ной соляной кислоте, в 3%-ной серной кислоте), растворяются в слабом растворе едкого натра без подогревания, в глицерине - при 300°С. Гемицеллюлозы клеточных оболочек нередко играют роль запасных веществ для растения, отлагающихся и затем расходующихся после гидролиза под действием фермента цитазы. Особенно мощны отложения гемицеллюлоз как запасных веществ в клеточных стенках эндосперма некоторых пальм (финиковой и др.). Семена пальмы Phytelephas macrocarpa крупные и очень твердые, их эндосперм используется как "растительная слоновая кость" для вытачивания пуговиц.
В целлюлозе клеточных оболочек мицеллярные пучки примыкают друг к другу с промежутками, поперечные размеры которых представляют величину порядка 10 мμ. Эти промежутки заполнены коллоидальными веществами, способными впитывать воду и при этом набухать. Такими веществами нередко являются пектиновые вещества (рис. 52).
Пектиновые вещества характеризуются хорошей растворимостью в щелочах после обработки кислотами и легкой окрашиваемостью красками - метиленовой синью (в сине-фиолетовый цвет), рутениевой красной, сафранином (в оранжево-желтый цвет).
Пектиновые вещества, хотя бы в небольших количествах, имеются в толще оболочек всех взрослых клеток; оказалось, что в оболочках, считавшихся ранее чисто целлюлозными (например, в хлопке), содержатся пектиновые вещества. В ранней стадии все клеточные оболочки высших растений состоят почти нацело из пектиновых веществ. Межклеточное вещество, как бы сцементировавшее оболочки смежных клеток, состоит обычно из пектиновых веществ, главным образом из пектата кальция.
Физико-химические свойства оболочки. Клеточные стенки представляют собой коллоиды в состоянии геля, т. е. они состоят из способных к набуханию, до максимума набухания, но нерастворимых веществ.
Клеточная оболочка отличается от других гелей, например от желатины или протоплазмы, своей анизотропностью.
Покажем на нескольких примерах, как проявляется анизотропия клеточных оболочек. Опытным объектом послужат целлюлозные лубяные волокна, например, льна или рами.
Если оставить льняные волокна в воде до максимального набухания, они увеличиваются в ширину на 20%, тогда как длина их увеличивается только на 0,1%. На рисунке 55 пунктиром изображена проекция воображаемых шариков из протоплазмы и оболочки; удлиненный эллипс есть проекция эллипсоида, образуемого набухшим шариком. Оси эллипса соответствуют направлению наибольшего и наименьшего набухания, они нанесены в виде двухконечных стрел. Получаемая таким образом картина
85
называется фигурой набухания. Эксцентричность эллипса указывает на степень анизотропии. Анизотропия набухания является важнейшей причиной разнообразных гигроскопических движений, наблюдаемых в мире растений.
Исследуя лучепреломление волокон льна, нашли, что показатель преломления в продольном направлении составляет 1,596, а в поперечном - 1,525. Анизотропия лучепреломления выражается в двойном лучепреломлении, известном у кристаллов. Численно оно равняется разности показателей преломления по двум направлениям и, следовательно, составляет для целлюлозных волокон 0,071; эта величина в 8 раз больше двойного преломления у кварца или гипса (0,009).
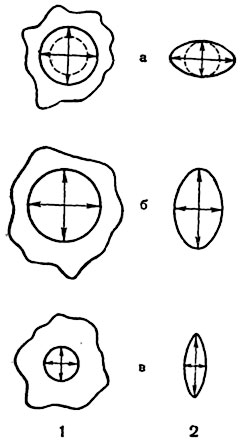
Рис. 55. Обозначение анизотропии:
1 - протоплазмы плазмодия; 2 - лубяного волокна (схематический продольный разрез через стенку волокна): а - анизотропия набухания, б - оптическая анизотропия (двойное лучепреломление); в - анизотропия прочности.
Способность преломлять и поглощать свет различна, в зависимости от направления волокна. Если окрасить волокна хлор-цинк-йодом и наблюдать препарат в линейно-поляризованном свете, идущем параллельно оси волокна, то оказывается, что свет поглощается почти целиком и волокно кажется глубоко черно-фиолетовым, тогда как перпендикулярно к оси волокна свет беспрепятственно проходит через клеточную стенку и волокно кажется бесцветным. Это явление двойного поглощения называют дихроизмом или плейохроизмом, если к двум упомянутым направлениям поглощения пространственно присоединяется еще третья ось поглощения.
Анизотропия физических свойств оболочки проявляется и в сопротивлении разрыву, и при исследовании эластичности клеточной стенки, и в ее тепло- и электропроводности. При воздействии на волокно параллельно и перпендикулярно его оси всегда оказывается большое различие в свойствах. Сопротивление разрыву в поперечном направлении составляет только 1/10 сопротивления в продольном направлении. Теплопроводность лубяного волокна липы по двум направлениям равна соответственно 5 и 3; коэффициент теплового расширения для волокон рами - 1 и 4.
Эта резкая анизотропия клеточной стенки тем замечательнее, что протоплазма, производящая клеточную стенку, не дает таких эффектов, связанных с направлением.
Образование стенки между двумя клетками. Обычно новая стенка появляется внутри существующей клетки, которая уже имеет оболочку. Каждая новая стенка между двумя клетками возникает при делении клеток. В поздней анафазе или телофазе в экваториальной плоскости фрагмопласта (стр. 53) образуются в ахроматиновых нитях зерновидные вздутия и одновременно с этим нити укорачиваются и исчезают; вещество каждой нити как бы стягивается в направлении от обоих ее концов к экваториальной плоскости. Зерновидные тельца, входя в соприкосновение и сливаясь, образуют срединную пластинку, или мембрану2. Если фрагмопласт доходит
86
до стенок клетки, то срединная пластинка формируется вся сразу. Если же экваториальное сечение фрагмопласта меньше поперечного сечения клетки, то срединная пластинка образуется постепенно: по периферии фрагмопласта возникают последовательно новые ахроматиновые нити до тех пор, пока фрагмопласт не войдет в соприкосновение с боковыми стенками клетки.
У некоторых семян при развитии эндосперма появление оболочки запаздывает; только после того, как ядро разделится несколько раз, появляются оболочки сразу у многих клеток (рис. 56). В некоторых случаях оболочка возникает не сразу поперек всей старой клетки, а постепенно, врастая от старой оболочки внутрь клетки.
Наблюдения над делением клеток показали, что скорость образования оболочки зависит от температуры (например, у спирогиры при 3-4° С образование оболочки заканчивалось через 14 часов, а при 12°С - через 45 минут, у традесканции при 10-11°С - через 2,5 часа, а при 40°С - через 35 минут.
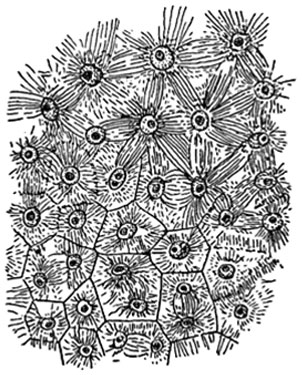
Рис. 56. Возникновение оболочек клеток в эндосперме резеды (Reseda).
Вверху - последовательное деление ядер, внизу - появление между ними сразу всех мембран.
Μембрана, или срединная пластинка, очень тонка, и очень скоро, уже в течение интеркинеза, если мы имеем дело с меристематической тканью, в перегородке между молодыми клетками вместо одного слоя можно различить три: средний из них - срединная пластинка, а боковые, обращенные в полости клеток и образовавшиеся позднее, точно повторяющие сетчатый рисунок срединной пластинки, - первичные слои клеточной оболочки. Срединная пластинка состоит из протопектина, пектина и пектиновых веществ; при образовании межклетников и при мацерации, как естественной, так и искусственной (стр. 103), происходит растворение срединной пластинки.
Первичная стенка в большей своей части состоит из протопектинов, гемицеллюлоз и других полисахаридов; содержание целлюлозы невелико (8-14%).
Довольно мелкие вначале меристематические клетки вырастают, их поверхность увеличивается соответственно увеличению объема иногда в несколько раз. Разрастание оболочки происходит как бы в одной плоскости: новые молекулы протопектина и целлюлозы внедряются между старыми молекулами (интусуссепция). Сразу после разрастания клетки,
87

Рис. 57. Расположение микрофибрилл целлюлозы (d = 100-300 A°) (электронный микроскоп, увел. 25 000).
а иногда одновременно с плоскостным ростом, который при этом может и продолжаться, начинается утолщение клеточной стенки. Утолщение происходит за счет наложения молекул целлюлозы - аппозиции.
Нередко, особенно при формировании прозенхимных клеток, происходит скользящий рост; клетки усиленно растут в одном (продольном) направлении, взаимно вклиниваясь заостренными концами; возможность такого роста, когда оболочки должны скользить одна по другой, обусловливается пластичностью межклетного вещества и молодых оболочек.
Через некоторое время после образования первичных клеточных оболочек, когда клетки перестают быть меристематическими, происходит значительное утолщение клеточной оболочки - вторичное утолщение. Вторичные слои формируются постепенно, только путем аппозиции, причем толщина слоев, направление слагающих их фибрилл и даже химизм несколько различны (рис. 57, 58). Эти слои по-разному
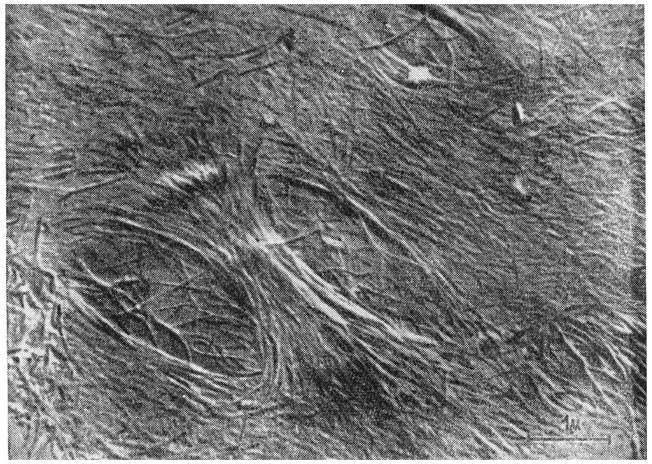
Рис. 58. Расположение микрофибрилл при образовании пор (электронный микроскоп).
88
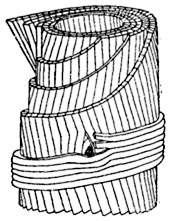
Рис. 59. Отпрепарированная (мысленно) часть клеточной оболочки лубяного волокна.
Оболочка построена из концентрических слоев толщиной каждый около 0,5μ; фибриллы в наружном слое образуют с направлением продольной оси волокна угол, близкий к 90°, а в других слоях - углы величиной от 0 до 30°.
преломляют свет и хорошо известны микроскопистам под названием слоистости клеточной оболочки (рис. 59).
В толще оболочки, особенно в длинных волокнистых клетках, удается наблюдать явление штриховатости. Штриховатость зависит от расположения фибрилл, составляющих оболочку (рис. 60). В слоях разных уровней системы штрихов или полос могут перекрещиваться, так как в разных слоях наклон фибрилл различен. Самый внешний, пограничный с первичными слоями слой вторичной оболочки часто обозначается как переходный, затем следует основная масса вторичной оболочки (рис. 61).
Между протоплазмой клетки и вторичным слоем клеточной оболочки находятся слои, как бы переходные между протоплазмой и вторичными слоями, которые часто называют третичными слоями (рис. 61, 62).
В некоторых случаях, во взрослых клетках, первичные оболочки и межклеточное вещество кажутся почти неразличимыми, и в этих случаях их все вместе также называют срединной пластинкой. На рисунках и схемах часто для простоты их изображают одной линией (рис. 61, 62).
Применение электронной микроскопии позволило изучить расположение микрофибрилл в оболочке. В первичной оболочке микрофибриллы располагаются более рыхло, и между ними остаются тончайшие капиллярные пространства. Во вторичных слоях фибриллы располагаются более плотно. Этим объясняется то, что первичные слои оболочки одревесневают сильнее вторичных слоев.
Утолщение оболочки. В молодых клетках все оболочки имеют одинаковую небольшую толщину и ровную гладкую поверхность. У вполне развитых клеток оболочка сформирована различно, что зависит от неравномерного разрастания в плоскости и в толщину. Строение и форма клетки стоят в тесной связи с отправлением клетки; часто по виду оболочки можно сказать, какова функция клетки (рис. 63, 64). Утолщение оболочки имеет, главным образом, механическое значение, и только у некоторых семян в сильно утолщенных оболочках откладывается запасное питательное вещество.

Рис. 60. Полосатость (штриховатость) клеточной оболочки склеренхимного волокна барвинка (Vinca sp.).
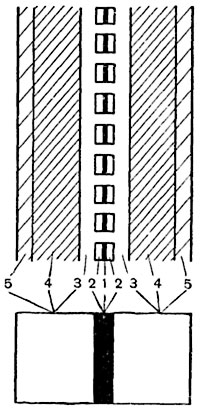
Рис. 61. Схема строения стенки между двумя клетками:
1 - срединная пластинка; 2 - первичные оболочки; 3 - переходные слои вторичной оболочки; 4 - основная масса вторичной оболочки; 5 - третичные слои.
89
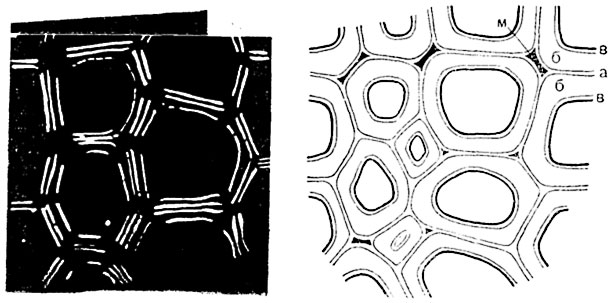
Рис. 62. Поперечный разрез древесины сосны.
Слева - в поляризованном свете: срединная пластинка и первичные оболочки заметны как две светлые линии с темным промежутком; внутренние светлые линии, окружающие полость клетки, соответствуют третичной оболочке. Справа - в обычном микроскопе: а - срединная пластинка и первичные оболочки, б - вторичная оболочка; в - третичная оболочка; м - межклетное пространство, возникающее после растворения срединной пластинки.
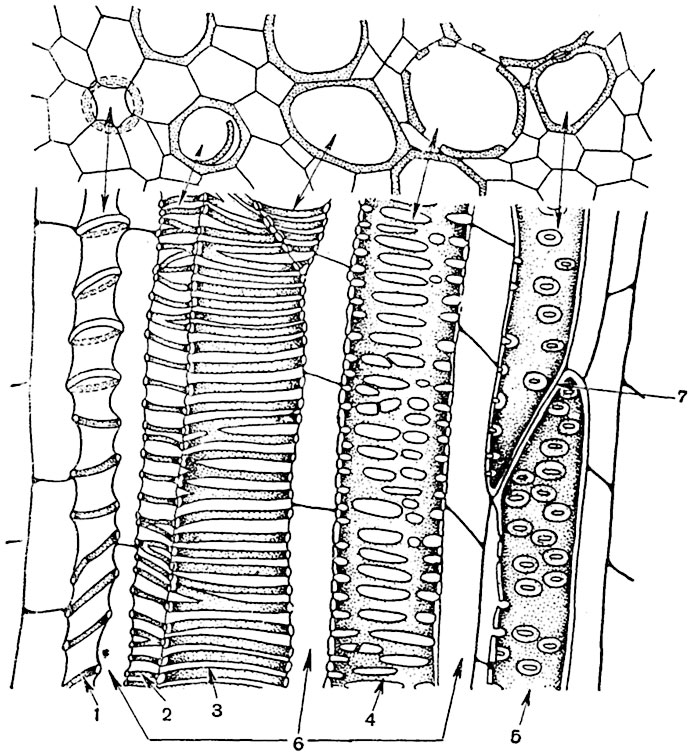
Рис. 63. Поперечный и продольный срезы молодого стебля кирказона:
1 - кольчатый сосуд; 2, 3 - спиральные сосуды; 4 - лестничный сосуд; 5 - пористый сосуд; 6 - клетки древесинной паренхимы; 7 - перфорация.
90
Утолщение оболочки не всегда идет равномерно вокруг всей клетки, чаще утолщению подвергаются лишь отдельные участки оболочки.
В исключительных случаях оболочка утолщается снаружи, это возможно только на свободной поверхности клеток, не соприкасающейся с оболочками соседних клеток. Наружное утолщение стенки бывает у клеток, которые развиваются внутри протоплазмы материнских клеток. Оно встречается на свободной поверхности клеток кожицы, на некоторых волосках, которые покрывают наземные органы или вдаются в воздушные полости внутри растения. Разнообразие наружных утолщений оболочек клеток
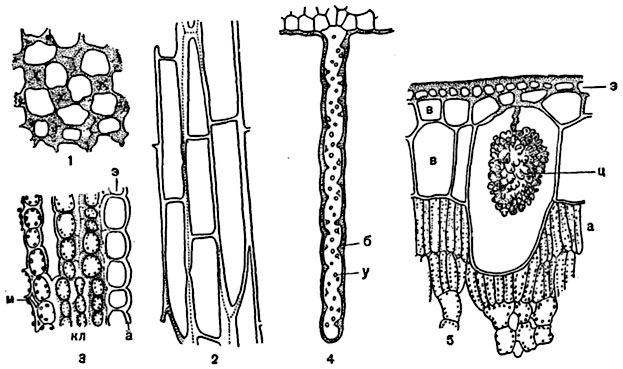
Рис. 64. Различные типы утолщения клеточной оболочки:
1, 2 - поперечный (1) и продольный (2) разрезы колленхимы листового черешка шалфея мускатного (Salvia sclarea) с уголковым утолщением стенок; 3 - колленхима с пластинчатым утолщением клеточных стенок, на поперечном разрезе черешка звездочки большой (Astrantia major): э - кожица с кутикулой (а), кл - клетки колленхимы с хлорофилловыми зернами, м - межклетники; 4 - продольно разрезанный ризоид печеночника (Marchantia polymorpha) с оболочкой, имеющей внутренние утолщения в виде бугорков, видимых в плане (у) и в оптическом сечении (б); 5 - часть поперечного разреза листа фикуса (Ficus elastica): э - кожица, в - водоносные слои, ц - цистолит, а - ассимиляционная паренхима.
пыльцы - в виде шипиков, гребешков, валиков - столь велико и в то же время настолько постоянно и характерно, что им можно пользоваться при классификации растений.
В неравномерном внутреннем утолщении стенки различают два случая: утолщенные части занимают меньшую часть всей оболочки; эти утолщенные места под микроскопом прежде всего обращают на себя внимание, и тогда говорят о скульптурных утолщениях оболочки. В другом случае утолщенные места занимают большую часть оболочки, и тогда бросаются в глаза места, оставшиеся в стенке неутолщенными, их называют порами. Конечно, в известной мере такое разделение произвольно и связано переходными образованиями.
Местные утолщения оболочки. Во многих случаях в оболочке ясно видно (иногда - после окрашивания оболочки) чередование утолщенных участков с тонкими; утолщенные участки видны, особенно при рассматривании стенки клетки сбоку или в плане, как отчетливый узор на
91
фоне более тонкого и потому более прозрачного общего наружного слоя оболочки. Основных скульптурных типов утолщения, при которых резко выделяются более толстые участки оболочки, имеется четыре: 1) кольчатое, 2) спиральное, 3) сетчатое и 4) лестничное. Местные утолщения оболочки имеют соответственно очертания: 1) параллельных колец, расположенных перпендикулярно к продольной оси клетки, 2) одной или нескольких лент, идущих по винтовой линии, или, как обычно выражаются, по спирали, 3) сетки, 4) лестницы с перекладинами. Встречаются утолщения промежуточных типов, например кольчато-спиральное (рис. 64), сетчато-лестничное. Утолщения упомянутых типов встречаются почти исключительно в стенках трахей (сосудов) или трахеид - продольных рядов или отдельных клеток, теряющих протопласт и служащих для передвижения в теле растения воды с растворенными в ней веществами. Очень распространен случай, когда утолщена большая часть поверхности оболочки, а тонкие места оболочки видны в стенке клетки как небольшие просвечивающие участки. Оболочку с утолщением такого рода называют пористой или точечной.
Поры представляют собой неутолщенные участки вторичной оболочки. Иногда замечаются более тонкие участки и в первичных оболочках, в этих случаях их называют первичными поровыми полями. Поры в стенках двух смежных клеток располагаются одна против другой, образуя пару пор. Пора может быть одиночной в том случае, если она находится в участке клеточной оболочки, граничащем с наружной средой или с межклетником.
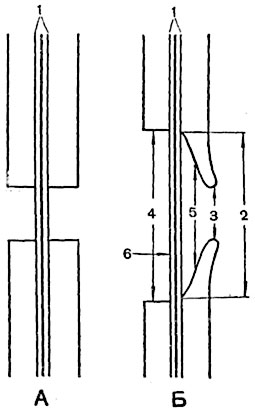
Рис. 65. Схема строения пор:
А - пара простых пор; Б - полуокаймленная пара пор; в Б слева пора паренхимной клетки, справа пора водопроводящей клетки; 1 - срединная пластинка и первичные оболочки; 2 - внутреннее отверстие поры; 3 - внешнее отверстие поры; 4 - канал поры; 5 - полость поры; 6 - замыкающая пленка поры.
Различают поры простые и окаймленные.
В простой поре общая полость клетки продолжается непосредственно в толщу стенки в виде полости поры до замыкающей пленки поры, являющейся неутолщенным участком клеточной оболочки (рис. 65, А). В толстостенных клетках полость поры представляет нередко цилиндрический (рис. 66), иногда изогнутый канал. Этот канал на одном конце сообщается внутренним отверстием с полостью клетки, а на другом конце упирается в замыкающую пленку поры. В очень толстостенных клетках полости двух-трех пор могут слиться в один канал, образуя ветвистую пору (рис. 66).
Полости (каналы) пор имеют поперечные сечения округлой, овальной, щелевидной формы. В паре овальных и щелевидных пор направления больших осей их поперечных сечений и их отверстий в плане перекрещиваются. Иногда перекрещивание видно в отдельно взятой поре; в этом случае форма поперечного сечения канала полости поры изменяется при переходе его из одного слоя стенки в другой.
Поры с цилиндрической полостью - округлые поры - образуются обычно в паренхимных клетках, а поры с щелевидной полостью -
92
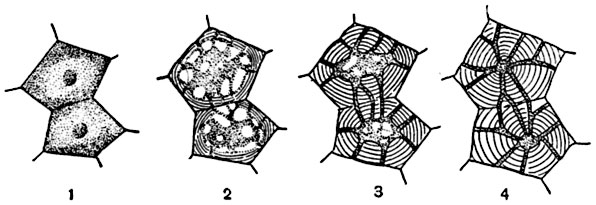
Рис. 66. Схема роста в толщину клеточной оболочки толстостенной клетки (1-4).
щелевидные поры - в прозенхимных клетках. Отверстия щелевидных пор располагаются косо по отношению к продольной оси стенки клетки. Размещаются такие поры обычно по винтовой линии, соответствующей направлению мицелл и фибрилл в оболочке. В том же направлении располагается и длинная ось отверстия поры.
Окаймленные поры, образующиеся в стенках сосудов, характеризуются тем, что полость поры суживается в направлении от замыкающей пленки к внутреннему отверстию поры. В полости окаймленной поры можно различить камеру поры и канал поры. Канал сообщается внешним отверстием с камерой поры, а внутренним - с клеточной полостью. Во многих случаях полость клетки примыкает непосредственно к отверстию камеры поры и пора почти не имеет канала (рис. 67).
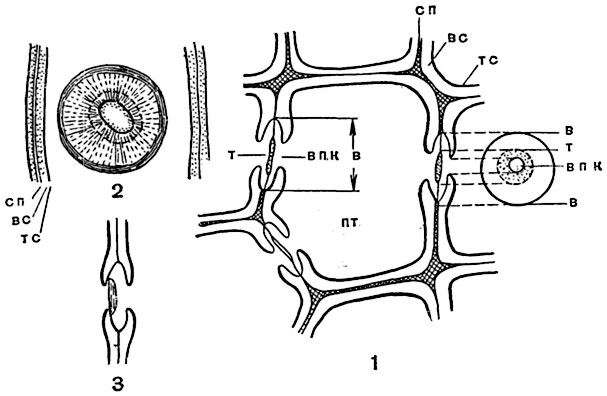
Рис. 67. Строение окаймленных пор в трахеидах древесины сосны (Pinus silvestris), несколько схематизировано:
1 - поперечный разрез одной трахеиды вместе со стенками смежных трахеид, примыкающими к ее оболочке; видны три окаймленные поры в разрезе, одна (справа) - в плане; 2 - часть продольного радиального разреза трахеиды при несколько большем увеличении; в поре, изображенной в плане, различаются очертания окаймления и отверстия канала поры; 3 - часть стенки трахеиды на продольном тангентальном разрезе через древесину; отверстие канала поры закрыто торусом; пт - клеточная полость трахеиды; сп - срединная пластинка стенки и первичные слои; вс - вторичные слои; тс - третичные целлюлозные слои оболочки; впк - внутреннее отверстие канала поры; в - наружное очертание окаймления и замыкающей пленки; т - очертание торуса.
93
Вторичная оболочка в окаймленных порах приподнимается над полостью поры, образуя как бы холмик с отверстием в середине (канал поры). В плане окаймленная пора в простейшем случае видна как две окружности: большая из них представляет наружное очертание окаймления, совпадающее с очертанием замыкающей пленки, а меньшая - очертание отверстия поры. Очертания окаймления и отверстий обеих пор одной пары в проекции совпадают.
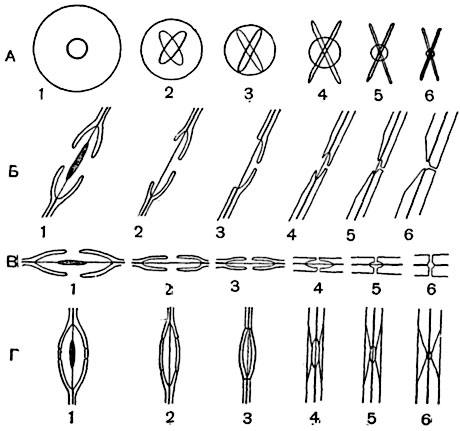
Рис. 68. Схема строения окаймленных пор шести типов:
А - вид в плане; Б, В - оптические разрезы: Б - по срединной плоскости отверстия порового канала, В - по срединной плоскости окаймления; Г - поры в профиль, на продольных срезах древесины. Пора типа 1 с торусом, прочие - без торуса.
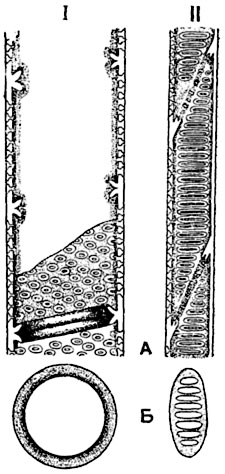
Рис. 69. Перфорации сосудов:
А - продольный срез; Б - поперечный срез; I - простая перфорация; II - лестничная перфорация.
Строение окаймленных пор может усложняться (у хвойных) тем, что срединная часть замыкающей пленки, утолщаясь, образует торус. При рассматривании таких пор в плане очертание торуса просвечивает в виде окружности, имеющей диаметр больший, нежели диаметр отверстия поры. На поперечных разрезах через пору по ее диаметру торус имеет очертание линзы (рис. 67). Встречаются (у сосен) окаймленные поры, в которых кольцевая окраина замыкающей пленки вокруг торуса имеет мелкие сквозные отверстия (перфорации); торус как бы подвешен в центре плоской сетки. Замыкающая пленка данной поры может изменять форму и положение; она или лежит в одной плоскости (находясь в среднем положении), или, искривляясь, прижимается изнутри к поверхности окаймления и замыкает отверстие поры (рис. 67, 3); в этом случае просачивание воды через пору прекращается. На это явление, как и на наличие иногда перфораций в замыкающих пленках пор, впервые обратил внимание Э. Руссов (1883)3.
94
В некоторых случаях строение окаймленной поры отличается от описанного типичного. Очертание внутреннего отверстия поры может иметь эллиптическую форму. При тесном расположении поры в очертании многоугольны. В сосудах и трахеидах с сильно утолщенными оболочками (как, например, в трахеидах поздней, осенней, древесины сосен) полость поры дифференцирована на камеру и длинный канал; канал щелевиден и имеет форму сплющенной воронки. Внутреннее отверстие канала поры имеет форму щели или весьма узкого прямоугольника с закругленными краями. Очертание внешнего отверстия близко по форме к окружности. Очертания щелевидных внутренних отверстий двух пор одной пары в плане часто перекрещиваются. Поры этого типа торуса иногда не имеют. Встречаются поры со строением, промежуточным между строением пор с округлым и щелевидным внутренним отверстием (рис. 68).
В лестничном утолщении тонкие участки оболочки являются не чем иным, как порами, и притом окаймленными; в плане каждая пора дает здесь картину двух узких овалов. Больший из них представляет очертание замыкающей пленки и вместе с тем внешнее очертание полости поры, а меньший - очертание внутреннего отверстия поры. Порами в широком смысле слова являются и тонкие участки оболочки, имеющей кольчатое, спиральное, сетчатое утолщение.
В паре пор не всегда обе супротивные поры одинаковы. Так, например, в парах пор в стенке, общей для двух соседних клеток, из которых одна выполняет функцию проведения воды, а другая служит хранилищем запасов органических веществ, пора со стороны первой клетки имеет окаймление, а пора на стороне второй его не имеет; такую пару пор называют полуокаймленной (рис. 65, Б).
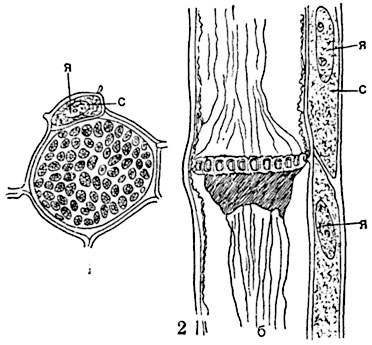
Рис. 70. Ситовидная трубка из стебля тыквы (Cucurbita pepo):
1 - ситовидная трубка с сопровождающей клеткой на поперечном срезе, в месте соединения двух члеников; хорошо видна ситовидная пластинка; 2 - ситовидная пластинка на продольном срезе: видны участки двух соседних трубок и их сопровождающих клеток; с - сопровождающие клетки; я - ядра сопровождающих клеток; б - протоплазма ситовидной трубки.
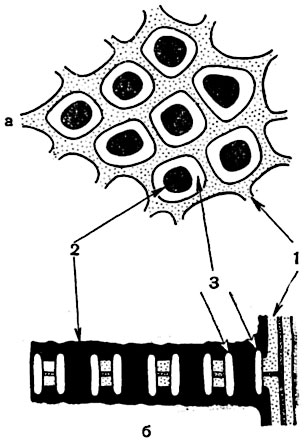
Рис. 71. Перфорация ситовидных трубок тыквы (Cucurbita pepo) при большом увеличении:
а - в плане; б - в продольном сечении: 1 - клеточная оболочка; 2 - протоплазма; 3 - каллеза.
95
Довольно крупные сквозные отверстия (перфорации) (рис. 69) встречаются в перегородке между члениками трахей, т. е. трубочек, по которым передвигается вода с раствором минеральных, а иногда и органических веществ (см. ниже). Более мелкие перфорации образуются в перегородках между члениками ситовидных трубок, по которым перемещается вода с растворами органических веществ (рис. 70-72).
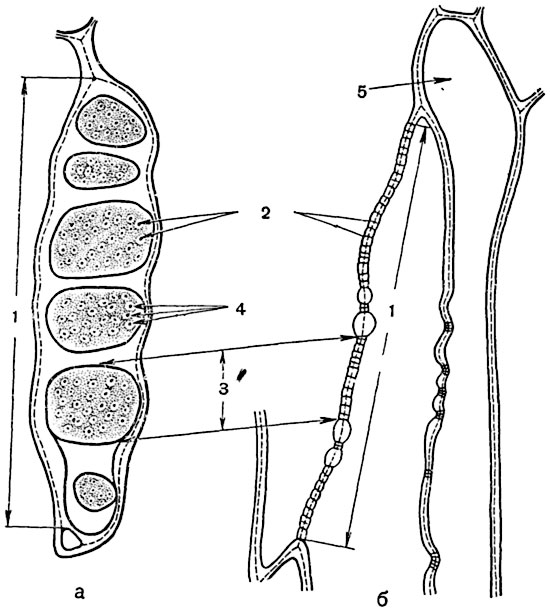
Рис. 72. Перфорация ситовидной трубки табака (Nicotiana):
а - в плане; б - в разрезе; 1 - сложная ситовидная пластинка; 2 - перфорации ситечек; 3 - отдельное ситечко; 4 - каллеза; 5 - паренхимная клетка.
Своеобразную форму утолщения клеточной стенки можно видеть в так называемых, цистолитах4, встречающихся у многих представителей семейств крапивных, тутовых, бурачниковых и некоторых других. Цистолиты образуются преимущественно в кожице листьев, реже в глубже лежащих клетках листа (а иногда стебля, корня).
Типичные цистолиты, как например цистолиты на верхней стороне листьев фикуса каучуконосного5, состоят из стержневидной ножки, прикрепляющейся одним концом к стенке клетки, а на другом конце несущей тело цистолита - мешковидное вздутие с многочисленными сосочковидными
96
выступами (рис. 64, 5). Тело цистолита слоисто; слои пронизываются в радиальном направлении тонкими трубчатыми канальцами, заканчивающимися сосочками. Каналец в каждом сосочке замкнут наружной тонкой оболочкой, облекающей тело цистолита.
Цистолит фикуса состоит из пропитанной кремнеземом ножки и гроздевидного тела, богатого углекислым кальцием.
По форме цистолиты весьма разнообразны: кроме гроздевидных, встречаются цистолиты с телом в виде веретена (рис. 73), шара (у крапивы), палицы и т. д.
Наблюдения и эксперименты показывают, что при накоплении в растении избытка извести цистолиты становятся вместилищами ее выделения, а в случае наступающего недостатка извести играют роль хранилищ запаса, который может быть снова использован растением.
Плазмодесмы. Кроме пор, в оболочках клеток есть образования другого рода - плазмодесмы - тончайшие нити, благодаря которым сообщается протоплазма соседних клеток. Плазмодесмы часто располагаются группами и могут пронизывать как всю утолщенную стенку, так и тонкие пленки в порах. Увидеть плазмодесмы без предварительной обработки или окраски трудно, их можно обнаружить окраской йодом в толще немного набухших стенок клеток чилибухи, хурмы, конского каштана, некоторых пальм, спаржи. Плазмодесмы связывают между собою протопласты всех клеток растения в одно живое целое, кроме того, по-видимому, служат для передачи по растению раздражений. По всей вероятности, плазмодесмы не могут принимать участия в перемещении веществ, так как капиллярное притяжение к стенкам чрезвычайно тонких плазмодесменных каналов будет настолько велико, что исключается возможность молярного передвижения по ним частиц протоплазмы. Плазмодесмы впервые замечены Э. Руссовым и И. Н. Горожанкиным, который описал плазмодесмы в 1879 г. (рис. 74).
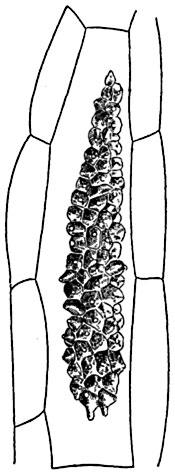
Рис. 73. Цистолит в листе Sanchezia sp. (из семейства акантовых).
Изменения в составе оболочки. Клеточная оболочка в процессе формирования нередко подвергается крупным изменениям в составе и тонкой структуре. В основном изменения могут быть сведены к следующим процессам: одревеснению, опробковению, кутинизации, ослизнению и минерализации.
Одревеснение. Одревеснение оболочки состоит в том, что в толще клеточной стенки откладывается лигнин. Лигнин, как и целлюлоза, состоит из углерода, водорода и кислорода, но относительное содержание углерода в нем значительно выше - 61-65% (в клетчатке - 44,5%). Он относится к соединениям ароматического ряда. Лигнин, выделенный из растения, представляет собой желтоватый аморфный порошок. Лигнин оптически изотропен, нерастворим в воде и в органических растворителях. Строение молекулы лигнина чрезвычайно сложно; на рисунке 75 представлен небольшой участок молекулы лигнина.
Одревесневшие оболочки, в отличие от состоящих почти нацело из целлюлозы, не растворяются в реактиве Швейцера, не
97
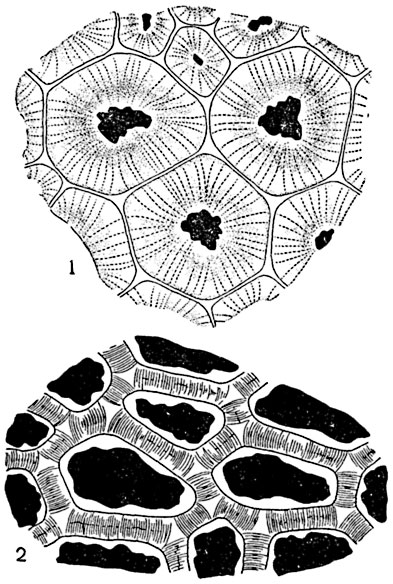
Рис. 74. Плазмодесмы в клеточных стенках:
1 - в эндосперме семени Sirychnos nux-vomica; 2 - в алейроновом слое зерновок кукурузы.
окрашиваются в синий цвет под действием реактивов, содержащих йод6.
Что касается цветных реакций на одревеснение, то их известно несколько десятков. Наиболее общеприняты следующие пробы на одревеснение: 1. Сернокислый анилин, особенно в присутствии небольшого количества серной кислоты, окрашивает одревесневшие оболочки в желтый цвет. 2. После обработки флороглюцином (в 1-5%-ном водном или спиртовом растворе), а затем концентрированной соляной кислотой появляется вишнево- или фиолетово-красное окрашивание одревесневших оболочек7.
3. По методу Мейле срез выдерживают в течение 5 минут в 1 %-ном растворе перманганата калия, затем промывают в течение 2-3 минут разведенной соляной кислотой; отмыв ее, действуют на препарат аммиаком (лучше всего, поднося препарат к открытому горлу склянки с водным раствором аммиака); одревесневшие оболочки становятся винно- или карминово-красными8.
Химический состав лигнина разных растений, по-видимому, различен. Древесина хвойных пород дает более яркое окрашивание, чем древесина лиственных пород при действии флороглюцина с соляной кислотой, а при реакции Мейле - наоборот.
Лигнин находится в клеточной оболочке, в промежутках между мицеллярными рядами целлюлозы. Если в препарате удалить из клеточных стенок только лигнин или только целлюлозу, то основной план строения клеточных стенок и ткани в целом остается не нарушенным. На основе этих данных и результатов исследований, произведенных с помощью поляризационного микроскопа и рентгеноустановок, создана теория, согласно которой одревесневшая оболочка в ее тонком строении аналогична железобетону; железной арматуре соответствует сетка из мицеллярных рядов целлюлозы, бетону - лигнин, заполняющий промежутки (ячейки) в сетке.
Одревеснение оболочек широко распространено в растительном царстве, за исключением низших растений и мохообразных; особенно богаты лигнином клетки древесины деревьев и кустарников. В редких случаях
98
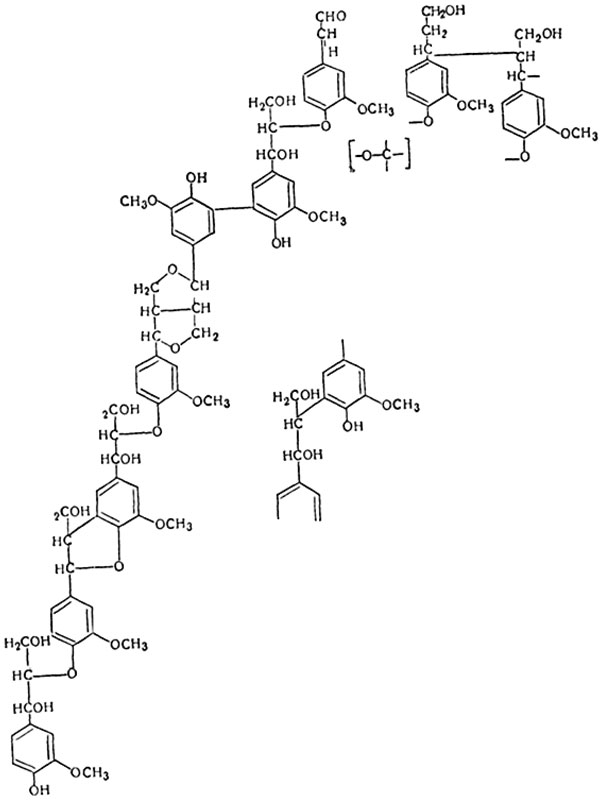
Рис. 75. Схема строения небольшого участка молекулы лигнина. Один из возможных вариантов строения.
происходит обратный процесс - удаление или разрушение лигнина, раздревеснение оболочек. Как пример можно привести явление, известное для плодов груши и айвы: в их мякоти появляются группы "каменистых клеток" (склереид) с сильно утолщенными одревесневшими стенками; при дозревании плода стенки склереид теряют лигнин и утоньшаются почти вдвое; после этого они окрашиваются при действии соответствующих реактивов как чисто целлюлозные.
В технике, в частности при фабрикации бумаги, искусственного шелка, раздревеснение соломы, древесины осуществляется кипячением измельченного сырья с едким натром под давлением выше атмосферного либо длительным кипячением с бисульфитом кальция.
Значение одревеснения оболочек для растений не вполне выяснено. При одревеснении повышаются твердость, плотность, калорийность,
99
лучепреломление; одревеснение понижает пластичность оболочек и прекращает их плоскостной рост9.
Лигнин, как и многие ароматические соединения, обладает консервирующими свойствами и придает, особенно мертвым клеткам, повышенную стойкость по отношению к разрушительному действию бактерий и грибов10.
Опробковение и кутинизация клеточных оболочек состоят в том, что в толще вторичной клеточной стенки (при опробковении) либо на ее наружной поверхности (при кутинизации, рис. 76) откладываются вещества, называемые соответственно суберинами и кутинами.
Суберины представляют собой глицерины феллоновой и пробковой кислот, нерастворимы в воде и спирте. Суберины стойки по отношению к концентрированной серной кислоте и реактиву Швейцера.
При подогревании среза пробки с едким кали до кипения образуются капли мыла - калийной соли феллоновой кислоты.
Суберин легко окрашивается некоторыми веществами, из которых назовем красную краску судан III (применяемый в смеси спирта с глицерином), свежеприготовленную спиртовую вытяжку хлорофилла и раствор альканнина (красного пигмента корней альканны красильной - Alcanna tinctoria из семейства бурачниковых) в спирту11.
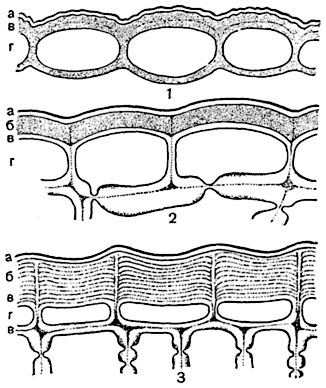
Рис. 76. Участки кожицы на поперечных разрезах:
1 - листа гвоздики перистолепестной (Dianthus plumarius); 2 - стебля Cereus triangularis (из семейства кактусовых); 3 - стебля Kleinia neriifolia (из семейства сложноцветных): а - кутикула, б - кутикулярные слои, в - целлюлозные слои, г - клеточная полость.
Хлор-цинк-йод, как и серная кислота с йодом, окрашивает суберины в буроватый цвет. Реакции на одревеснение отрицательны.
Пропитанные суберином (опробковевшие) оболочки клеток непроницаемы для воды, паров и газов.
Опробковению подвергаются преимущественно клетки покровной ткани - пробки. Часто опробковевают стенки клеток, расположенных вблизи поврежденного участка (поранения, погрызы, морозобоины, бактериальные заболевания и др.); благодаря этому поврежденный участок изолируется от остальных здоровых тканей (частей) растения.
Кутины представляют собой смесь различных воскоподобных веществ. По отношению к серной кислоте кутины более стойки, чем суберины. Едкие щелочи действуют на кутины более слабо и медленно. Кутин
100
отлагается обычно в виде пленки, называемой кутикулой, на наружной поверхности клеток кожицы листьев и стебля; нередко образуются, кроме того, в целлюлозной толще наружной стенки клеток кожицы тонкие прослойки кутина - кутикулярные слои (рис. 76). Благодаря стойкости кутина кутикула хорошо сохраняется в палеонтологических остатках.
В наружном слое оболочки спор, пыльцевых зерен семенных растений содержатся спорополленины, аналогичные суберинам и кутинам и отличающиеся от них наличием жирных кислот и жиров, еще более высокомолекулярных. Спорополленины наиболее стойки к внешним воздействиям. Благодаря им споры и пыльцевые зерна чрезвычайно хорошо сохраняются в различных геологических отложениях.
Резкой границы между тремя вышеупомянутыми типами веществ нет. Различают их главным образом по стойкости к реактивам: при действии
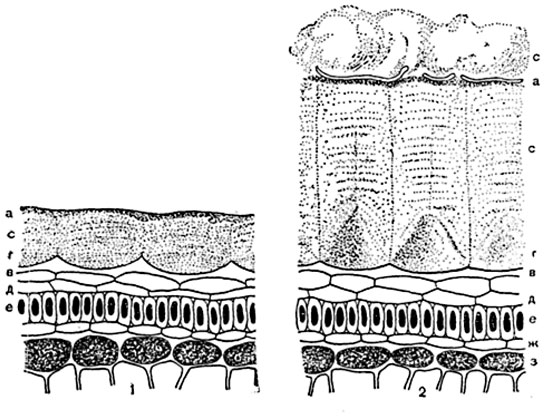
Рис. 77. Поверхностные слои семени льна (Linum usitatissimum) на поперечном разрезе:
1 - сухого семени (препарат в канадском бальзаме); 2 - семени, набухшего в воде; α - кутикула; с - ослизненный слой наружной стенки кожицы; г - клеточные полости этих клеток; в - внутренняя стенка их; д - два слоя тонкостенных клеток; е - "палисадный слой" - ряд столбчатых толстостенных клеток; ж - слой низких тонкостенных клеток; з - слой клеток с бурым содержимым.
5%-ной горячей едкой щелочью суберины омыляются быстро, кутины - медленно, спорополленины вовсе не омыляются; деполимеризуются при обработке кипящим глицерином: суберины - быстро, кутины - медленно, спорополленины - медленно и слабо.
Ослизнение оболочек. При ослизнении оболочки содержащиеся в ней целлюлоза и пектиновые вещества превращаются в слизи и камеди - вещества, пока еще с химической стороны мало изученные, главным образом ввиду трудности получения их в химически чистом виде. В камеди вишневого дерева установлено наличие гемицеллюлоз типа арабанов (C5H8O4) n.
Нормальное ослизнение оболочек представляет приспособительное явление; так, при ослизнении слои наружных стенок кожицы у семян (льна и других растений, рис. 77), весной набухают от воды и, прорывая кутикулу, входят в соприкосновение с почвой. Благодаря клейкости слизь закрепляет семя на влажном месте и, поглощая воду почвы и атмосферных
101
осадков, обеспечивает прорастающее семя водой. Слизь, образующаяся в клетках кожицы листьев некоторых растений пустынь и полупустынь (например, по Б. А. Келлеру, у Rosa persica), образует покров, защищающий лист от перегревания, хранящий запас воды и даже, быть может, адсорбирующий водяные пары из воздуха12.
Минерализация оболочек имеет место в небольшой мере у всех клеток. В некоторых случаях минеральные вещества отлагаются в значительных количествах в толще оболочки или на ее поверхности, а иногда в особых выростах оболочки; отложения могут быть аморфными или кристаллическими. Наиболее распространены отложения кремнезема и солей кальция.
Кремнеземом инкрустируются оболочки клеток кожицы стеблей и листьев хвощей, злаков (рис. 78), осоковых; кремнеземом сильно пропитаны наружные и слабее боковые стенки кожицы листьев многих древесных растений (каркаса - Celtis, магнолии крупноцветной - Magnolia grandiflora) и трав (ясменников - Asperula)13.
Рис. 78. Сподограмма (кремневый скелет, полученный после обработки препарата соляной кислотой с последующим прокаливанием) участка кожицы бамбука (Bambusa sp.).
Вид в плане. Видны пары замыкающих клеток устьиц и клетки кожицы с извилистыми стенками; некоторые из них сплошь заполнены кремнеземом.
Для точного установления наличия кремнезема в оболочках препарат обрабатывают соляной и азотной кислотами и затем после промывки водой действуют плавиковой кислотой, растворяющей кремнезем14 (рис. 78).
Кальций встречается в клеточных оболочках в виде углекислого и щавелевокислого кальция. У многих растений углекислый кальций отлагается на поверхности кожицы, осаждаясь из выделений водных устьиц. Углекислый кальций вместе с кремнеземом встречается в клеточных стенках жгучих волосков (у крапив), в выростах клеточных стенок, называемых цистолитами (рис. 64, 73). Щавелевокислый кальций откладывается в виде кристаллов в клеточных оболочках некоторых растений (например, у вельвиччии15, в толстостенных ветвистых "спикулярных" клетках).
102
1 У грибов клеточная оболочка значительно отличается от клетчатки: не дает реакции с хлор-цинк-йодом, не растворяется в швейцеревом реактиве; в составе ее отмечен хитин. У лишайников в составе оболочки указано вещество, близкое к амилоидному видоизменению клетчатки, - лихенин.
2 У низших растений при делении клеток, как одноядерных, так и многоядерных, образование поперечной перегородки не связано с ахроматиновым аппаратом: оно начинается в то время, когда ахроматиновая фигура уже исчезла. Новая перегородка формируется центростремительно: в протоплазме подле боковых стенок клетки появляется кольцевая перетяжка, заполняющаяся твердым веществом новой перегородки. Перетяжка плазмы завершается в центре, и новая клеточная стенка становится сплошной пластинкой, перегораживающей материнскую клетку на две дочерние. У водорослей с одноядерными клетками перегородка закладывается между двумя ядрами в экваториальной плоскости кариокинеза, приведшего к образованию этих ядер. В многоядерных клетках, характерных для многих грибов и некоторых водорослей, ядра делятся обычно независимо друг от друга и формирование перегородки при делении клеток не стоит в связи с кариокинезом.
3 Эдмунд Руссов (1841-1897), крупный исследователь, был профессором ботаники университета в Юрьеве (Дерпте).
4 От греческих "кюстис" - пузырь, "литое" - камень.
5 Ficus elastica - тропическое растение из семейства тутовых, обычное у нас в комнатной культуре.
6 Одревесневшие оболочки окрашиваются от йода и серной кислоты в желтый цвет.
7 По предложению А. Н. Бояркина, HCl заменяют 25%-ной H2SO4. Этим избегается вредное действие паров HCl на микроскоп, но окрашивание при этом происходит значительно медленнее.
8 Соляная кислота может быть заменена серной или фосфорной, раствор аммиака - едкой щелочью или раствором соды.
9 Клетки с одревесневшими оболочками могут не только оставаться живыми в течение десятков лет, но иногда даже расти и делиться; в этих случаях, вероятно, происходит временное раздревеснение.
10 Однако многие грибы (трутовики) используют с помощью фермента гадролазы лигнин клеточных стенок.
11 Надо иметь в виду, что судан III окрашивает также жиры, смолы и воск; окраска от хлорофилла очень кратковременна, а альканнин окрашивает, хотя медленно, одревесневшие оболочки.
12 Путем ослизнения оболочек образуются покровные футляры у многих водорослей.
13 Особенно богаты кремнеземом створки диатомовых водорослей.
14 При этом должны, конечно, соблюдаться технические условия, необходимые при работе с плавиковой кислотой.
15 Welwitschia mirabilis - многолетнее растение из класса оболочкосеменных, характерное для почти бездождных пустынь Юго-Западной Африки.