

ОПЛОДОТВОРЕНИЕ
Между опылением и оплодотворением, т. е. слиянием гамет, проходит больший или меньший промежуток времени. У каучуконоса кок-сагыза он составляет всего 15-30 минут, у хлопчатника - 18-20 часов, у некоторых - несколько дней, недель, месяцев и даже год.
Пылинка, попав на рыльце, начинает прорастать (рис. 316); содержимое ее, одетое интиной, выпячивается через поры в экзине (см. стр. 336) и образует пыльцевую трубочку, которая постепенно удлиняется и растет по каналу столбика, если он есть, или по особой проводящей ткани в столбике, направляясь в завязь. Необходимые для ее питания и роста вещества пыльцевая трубочка получает, по-видимому, в значительной степени не из запасов пылинки, а из тканей столбика. Направление роста пыльцевой трубочки тоже обусловливается воздействием каких-то веществ ткани столбика и семяпочек. Содержимое пылинки - протоплазма с вегетативным ядром и генеративная клетка - переходят в растущий кончик пыльцевой трубочки; у некоторых растений, впрочем, вегетативное ядро остается в пылинке. Генеративная клетка иногда еще в пыльнике, чаще же в пыльцевой трубочке, делится на две. Раньше обычно принимали, что границы этих дочерних генеративных клеток исчезают, протоплазма их смешивается с протоплазмой вегетативной клетки и хорошо обособленными являются лишь два генеративных ядра, которые называют также спермиями1 (бесплазменными спермиями). В настоящее время установлено, что спермии представляют собой не одни лишь ядра, а что/ генеративные ядра окружены тонким слоем принадлежащей им протоплазмы и что, следовательно, в пыльцевой трубочке находятся не бесплазменные спермии, а спермии-клетки, имеющие обыкновенно вытянутую форму. Эти наблюдения имеют большое принципиальное значение: они доказывают, что в оплодотворении, производимом спермиями, с мужской стороны участвует не одно лишь ядро, а ядро и протоплазма.
В протоплазме пыльцевой трубочки различимы бывают у растущего конца ее вегетативное ядро, которое постепенно разрушается, а позади
371
него два спермия, отличающиеся большей компактностью и вытянутой формой (рис. 317). Достигнув завязи, пыльцевая трубочка растет по особой, проводящей ткани ее, достигает семяпочки и проникает в нее через пыльцевход. Здесь она или прямо встречает зародышевый мешок, или проникает к нему через разрыхленные, прикрывающие его клетки. Оболочка зародышевого мешка, соприкасающаяся с концом пыльцевой трубочки, растворяется. Войдя в зародышевый мешок, пыльцевая трубочка растет между синергидами или между одной из синергид и стенкой зародышевого мешка по
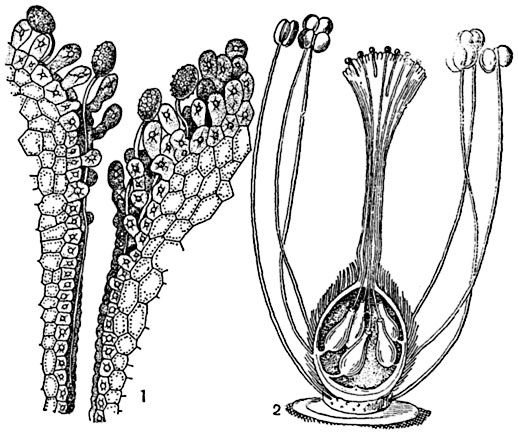
Рис. 316. Прорастание пыльцы и рост пыльцевых трубочек:
1 - прорастание пыльцы на рыльце; 2 - схематическое изображение роста пыльцевых трубочек к семяпочкам (прямым).
направлению к яйцеклетке. Оболочка на кончике пыльцевой трубочки разрывается, из нее выходят два спермия, которые направляются: один - к ядру яйцеклетки, другой - ко вторичному ядру в центральной клетке зародышевого мешка и сливаются с ними (рис. 317). Ряд исследователей описывают при этом активные движения спермиев2.
Таким образом, происходит так называемое двойное оплодотворение - особенность покрытосеменных, не наблюдаемая у голосеменных. Из оплодотворенной яйцеклетки развивается зародыш, из центральной клетки зародышевого мешка - эндосперм, идущий на питание зародыша; синергиды и антиподы отмирают. Двойное оплодотворение впервые было открыто выдающимся русским ботаником С. Г. Навашиным в 1898 г.
У некоторых растений пыльцевая трубочка проникает в семяпочку не через пыльцевход, а через халацу (халацогамия) или сбоку через покровы (мезогамия, от греческих "мезос" - средний, "гамео" - вступаю в брак). Оба последних способа возникли у различных семейств в процессе эволюции из первого (так называемой порогамии).
372
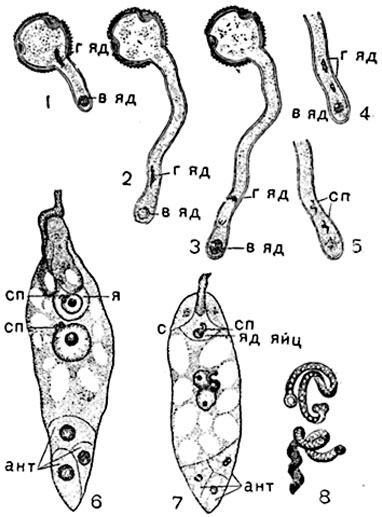
Рис. 317. Оплодотворение у покрытосеменных:
1-5 - рост пыльцевой трубочки и формирование спермиев (схемат.); 6 - двойное оплодотворение у лилии (Lilium marfagon); 7 - двойное оплодотворение у подсолнечника (Helianthus annuus); 8 - спермии у него же; г яд - генеративное ядро; в яд - вегетативное ядро; сп - спермии; с - синергиды; я - яйцеклетка; яд яиц - ядро яйцеклетки; ант - антиподы.
В ряде работ некоторое значение придается так называемому избирательному оплодотворению у культурных цветковых растений. На рыльце попадает обычно много пылинок, могущих оплодотворить яйцеклетки в семяпочках. Но оплодотворение, по данным этих работ, производят не любые пылинки даже того же вида или того же сорта, случайно опередившие другие, а те пылинки, оплодотворение которыми даст наилучшие результаты, явится биологически наиболее выгодным. Эту способность избирательного оплодотворения в настоящее время широко используют в СССР при скрещивании культурных растений. Выявлен еще ряд интересных подробностей в опылении многих культурных растений. И. В. Мичурин при скрещивании двух плохо скрещивающихся видов прибавлял к пыльце отцовского растения в небольшом количестве пыльцу материнского вида, и это способствовало успеху опыления чужой пыльцой. Количество пыльцы, взятой для опыления, тоже имеет значение для успешности опыления. У некоторых растений одиночно прорастающие пылинки дают пыльцевые трубки, отстающие в росте от тех, которые прорастают группами или даже вовсе не прорастающие. Для успешности опыления количество пылинок должно во много раз превышать число опыляемых семяпочек. Возможно, что в этих случаях известную роль играют какие-то вещества, выделяемые пыльцой и необходимые для начальных стадий образования плодов.
373
1 От греческого "сперма" - семя, так как они служат для оплодотворения, как мужское "семя" у животных.
2 У некоторых видов второй спермий сливается одновременно с общими, еще не слившимися полярными ядрами; у некоторых спермий сливается сначала с одним полярным ядром и затем с ними сливается второе полярное ядро.