

2. Мейоз, или редукционное деление
Мейоз характеризуется уменьшением числа хромосом вдвое. С точки зрения смены ядерных фаз в цикле развития растений редукционное деление представляет процесс, как бы противоположный оплодотворению. Как известно, сущность оплодотворения заключается в слиянии двух половых клеток (гамет), причем получается новая клетка - зигота, представляющая результат слияния их протоплазм и клеточных ядер.
Образовавшаяся зигота характеризуется двойным числом хромосом (диплоидное ядро) по сравнению с тем числом, которое характеризует ядра гамет (гаплоидные ядра). Таким образом, при каждом оплодотворении потенциальное число хромосом в ядре удваивается. Понятно, что при этих условиях в ряде следующих друг за другом поколений, происходящих половым путем, число хромосом в ядре увеличивалось бы до бесконечности. Этого не происходит, так как каждому удвоению числа хромосом при оплодотворении противопоставляется уменьшение его вдвое в процессе мейоза. Мейоз имеется у каждого организма с половым циклом развития. Что же касается места его в онтогенезе, то оно различно у разных организмов. У животных мейоз происходит перед самым оплодотворением, точнее при формировании гамет, а у цветковых растений - незадолго до оплодотворения - при развитии микроспор (пыльцы) и мегаспор, дающих начало зародышевым мешкам. У архегониальных растений мейоз значительно отодвинут от оплодотворения и также происходит при формировании спор, а у многих низших растений он следует непосредственно за оплодотворением при делении зиготы. Благодаря этому создаются различные соотношения ядерных фаз в цикле развития разных организмов. У животных преобладает диплоидная фаза (от зиготы до образования гамет взрослой особью), а гаплоидная представлена только гаметами. У цветковых растений также преобладает диплоидная фаза, но и гаплоидная представлена несколькими клетками (от пыльцевого зерна до образования спермиев в пыльцевой трубке и от мегаспоры - материнской клетки зародышевого мешка - до образования в нем яйцеклетки). У папоротников также преобладает диплоидная фаза, но значительно развита и гаплоидная. Последняя представлена здесь заростком, развивающимся из споры и дающим половые органы с гаметами в них, диплоидная же фаза представлена самим папоротником, развивающимся из зиготы и дающим споры, при образовании которых происходит мейоз. Таким образом, у папоротников диплоидная и гаплоидная фазы представлены самостоятельно живущими и различными по внешности поколениями: гаплоидная - половым, или гаметофитом, в
54
виде заростка, диплоидная - бесполым, или спорофитом, в виде самого папоротника. Эти поколения правильно чередуются в общем цикле развития (гаметофит→спорофит→гаметофит→спорофит и т. д.) (см. стр. 301). У многих низших растений мейоз имеет место непосредственно после оплодотворения, при первом делении ядра зиготы, развивающейся в новую особь с гаметами на ней. Таким образом, в этом случае вся вегетативная жизнь протекает в гаплоидной фазе и диплоидна только зигота, имеющая притом характер покоящейся клетки.
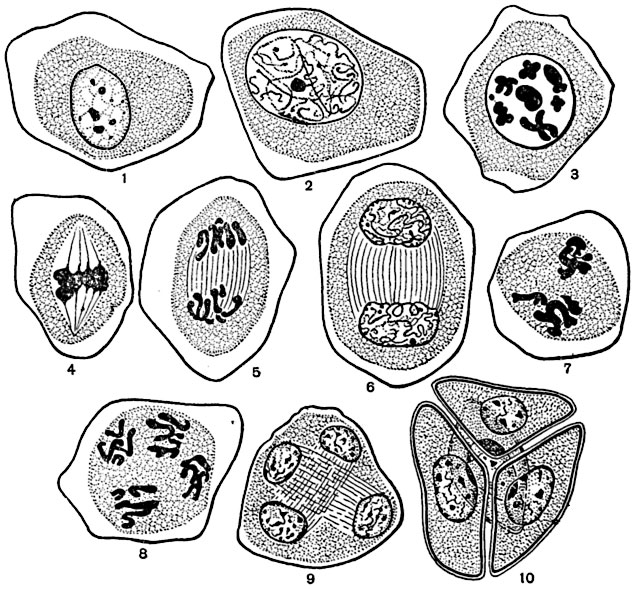
Рис. 25. Мейоз в материнских клетках пыльцы эремуруса:
1-6 - гетеротипное деление; 7-10 - гомеотипное деление; 1 - интерфаза; 2 - ранняя профаза (лептонема); 3 - диакинез; 4 - ранняя анафаза; 5 - поздняя анафаза; 6 - телофаза, переходящая в профазу гомеотипного деления; 7 - метафаза; 8 - анафаза; 9 - поздняя телофаза; 10 - сформированная тетрада микроспор.
В мейозе число хромосом уменьшается вдвое. Мейоз состоит из двух быстро следующих друг за другом делений ядра. Из этих делений первое называется гетеротипным (от греческого "гетерос" - различный), второе - гомеотипным (от греческого "гомойос" - одинаковый). Эти названия даны потому, что в первом делении происходит существенная перестройка хромосомного аппарата ядра, в результате чего два дочерних ядра оказываются различными и гаплоидными, а во втором делении, сходном с обычным митозом, образуются одинаковые дочерние ядра, в данном случае тоже гаплоидные (рис. 25, 26).
В мейозе, как и в митозе, различают профазу, метафазу, анафазу и телофазу.
55
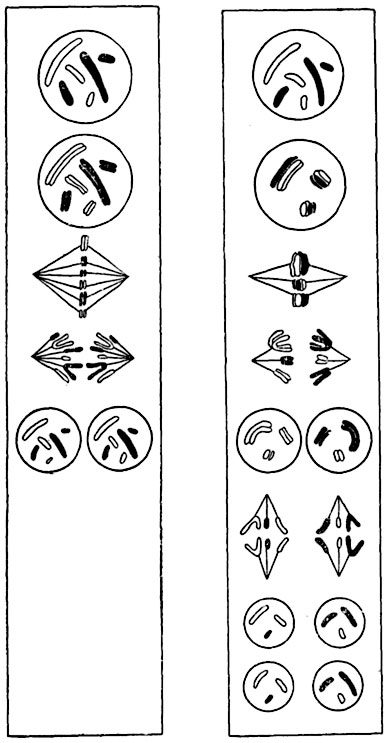
Рис. 26. Схематическое сопоставление эквационного деления ядра (левый столбец) с редукционным (правый столбец).

Рис. 27. Схема образования хиазм.
Слева - две гомологичные хромосомы, состоящие каждая из двух хроматид, образовали хиазму (перекрест); посредине - две хроматиды. обменялись участками; справа - четыре хроматиды, из которых две обменялись участками.
Профаза гетеротипного деления - наиболее длительная и сложная фаза, так как здесь происходит подготовка к редукции числа хромосом. Профазу гетеротипного деления подразделяют на несколько стадий.
В ранней профазе - лептонеме - хромосомы заметны в виде отдельных очень тонких хроматиновых нитей (рис. 25, 2). Известно, что на этой стадии число нитей равно диплоидному числу хромосом.
Немного позднее, в зигонеме, хроматиновые нити сближаются попарно, соединяясь во многих точках. При этом, по-видимому, происходит не случайное, а выборочное сближение: сближаются только гомологичные хромосомы, т. е. одинаковые по форме и происходящие от разных гаплоидных ядер.
Еще позднее, в пахинеме, нити (гомологичные хромосомы) соединяются (конъюгируют) по всей длине и заметно укорачиваются. Двойственная природа образовавшихся хромосом плохо заметна. Общее число двойных хромосом гаплоидно.
Далее, в диплонеме, каждая гомологичная хромосома расщепляется на две хроматиды, которые остаются соединенными лишь в отдельных точках. Таким образом, каждая двойная хромосома содержит четыре хроматиды.
В конце профазы - диакинезе - двойные хромосомы очень сильно укорачиваются, утолщаются и располагаются по периферии ядра (рис. 25, 3). В диакинезе очень легко подсчитать число хромосом (бивалентов) и точно установить, что оно гаплоидно.
Довольно часто,
56
по-видимому как результат фиксации, в ранних стадиях профазы хромосомы собираются в более или менее плотный комок, сдвигающийся к одной стороне ядра. Это явление называется синапсисом или синизезисом.
В метафазе появляется ахроматиновое веретено, исчезают ядерная оболочка и ядрышки, которые, как и в митозе, не принимали непосредственного участия в указанных перегруппировках. В поздней метафазе хромосомы в гаплоидном числе слагаются в ядерную пластинку. Они здесь заметно отличаются по форме от типичных хромосом митоза (в общем более короткие и толстые) и представляют биваленты, т. е. пары сближенных гомологичных хромосом. В метафазе каждая хромосома сначала бивалентна, затем она расщепляется вдоль, так что в результате получается образование из четырех элементов, разделенных продольными щелями в двух взаимно перпендикулярных направлениях: одна щель, называемая редукционной, отделяет гомологичные хромосомы друг от друга, а другая, называемая эквационной, отделяет их хроматиды, т. е. половинки.
В анафазе происходит расхождение гомологичных хромосом, т. е. разъединение их по редукционной щели (истинная редукция), но в это время каждая из расходящихся хромосом уже состоит из двух сформированных хроматид (рис. 25, 4, 5).
За гетеротипным сейчас же следует гомеотипное деление. Поэтому телофаза первого сильно сокращается, как сокращается и профаза второго. Телофаза непосредственно переходит в профазу гомеотипного деления (рис. 25, 6). В анафазе хромосомы расходятся по эквационной щели (рис. 25, 8).
При гетеротипном делении гомологичные хромосомы распределяются случайно и хромосомы каждого бивалента могут с равной вероятностью отойти к одному или другому полюсу делящегося ядра.
Следует отметить, что в профазах мейоза во время конъюгации гомологичных хромосом они переплетаются друг с другом и в одном или нескольких участках перекрещивания (кроссинговера) может происходить их слипание. При последующем расхождении хромосом половинки их (хроматиды) могут обменяться, и в этом случае хромосома будет составлена из участков разных хромосом. Этот обмен происходит в местах соприкосновения - хиазмах (от греческого "хиазмос" - перекрест, расположение в виде буквы "х") (рис. 27).
Особо большое внимание в цитогенетических работах по изучению мейоза было обращено на вопросы конъюгации гомологичных хромосом и возможный при этом обмен их частей.
Вопросы образования хиазм считаются важными при цитогенетических работах по изучению мейоза.
По окончании мейоза происходит образование новых клеток. Иногда перегородки образуются последовательно после первого и второго делений, а иногда каждая из четырех новых клеток образует оболочку внутри старой материнской клетки только после второго деления (рис. 25).
Такое образование клеток - свободное образование клеток - происходит при образовании спор в спорангиях у ряда споровых растений (см. в разделе "Морфология", стр. 344), в том числе аскоспор сумчатых грибов (их спорангии называют сумками), и в некоторых случаях при образовании пыльцевых зерен в пыльниках цветковых растений. В многоядерном протопласте вокруг ядер обособляются участки, которые облекаются твердой оболочкой.
Каждая клетка окружена эпиплазмой, массой протоплазмы материнской клетки, расходующейся на образование оболочек дочерних клеток. Благодаря этому в ряде случаев на оболочке образуются скульптурные утолщения.
57